C2005/F2401 '10-- Lecture 15
-- Last Edited: 11/04/10 02:35 PM
© Copyright 2010
Handouts: 15A --
Induction vs Repression; Repression vs
Feedback Inhibition &
15B --
Operons
I. Summary of 'How RNA makes protein'. See notes for lecture 14, topic IV.
II. Mutations. See notes for lecture 14 -- Topic V.
III. Intro to Regulation in Prokaryotes (See handout 15A)
A. Why regulation of enzyme synthesis is reasonable and/or necessary -- consider some typical enzymes -- glycolytic enzymes, beta-galactosidase (needed to breakdown and metabolize lactose = dimer of glucose and galactose), and trptophan synthetase (needed to synthesize trp). (See Becker 23-1 & 23-2.) When are these enzymes needed?
1. Glycolytic enzymes -- always needed
2. Beta-galactosidase -- only needed if lactose present (and needs to be broken down); enzyme level should be low until lactose added to medium.
3. TS (trp synthetase) -- only needed if trp low or absent (then trp must be synthesized in order to make proteins) -- enzyme level should be high until trp added to medium.
4. Why not make all enzymes all the time (even if not needed)? Enzyme synthesis uses a lot of energy.
B. The Phenomena -- Are enzymes (like those above) actually made only when they are needed? Graphs on handout 15A show what happens to level of appropriate enzyme if you add, or take away, the appropriate small molecule, namely lactose (lac) or tryptophan (trp).
1. Example of Induction -- Lactose (small molecule) = inducer = signal to turn on synthesis of appropriate enzyme ; synthesis of beta-galactosidase (enzyme) is called inducible; phenomenon is known as induction. See also Sadava fig. 16. 8 (13.16)
2. Example of Repression -- tryptophan (small molecule) = co-repressor = signal to turn off synthesis of appropriate enzyme ; synthesis of trp synthetase (enzyme) is called repressible; phenomenon is known as repression.
3. Constitutive synthesis -- Synthesis of some proteins, such as enzymes of glycolysis, is called constitutive = synthesis of enzymes is "on" at all times.
C. Summary of Terminology = italicized terms above. See table in middle of Handout 15A.
Regulation is covered in problem set 12. To review the material in parts A-C, see Problem 12-1, parts A & B.
D. Comparison of Repression to feedback. Why do you need both types of regulation? Factors to consider:
Overall, have coarse control (repression/induction) vs fine control (inhibition/activation). See chart and picture on bottom half of handout 15A. See also Sadava fig. 16.9 (13.17). Note: Enzyme activation and induction can be compared in a similar way -- Activation increases enzyme activity while induction turns on enzyme synthesis
Today's lecture will focus on induction; we will go over the mechanism of repression in detail next time. Wait to do the problems on repression and/or repression vs. feedback until next time.
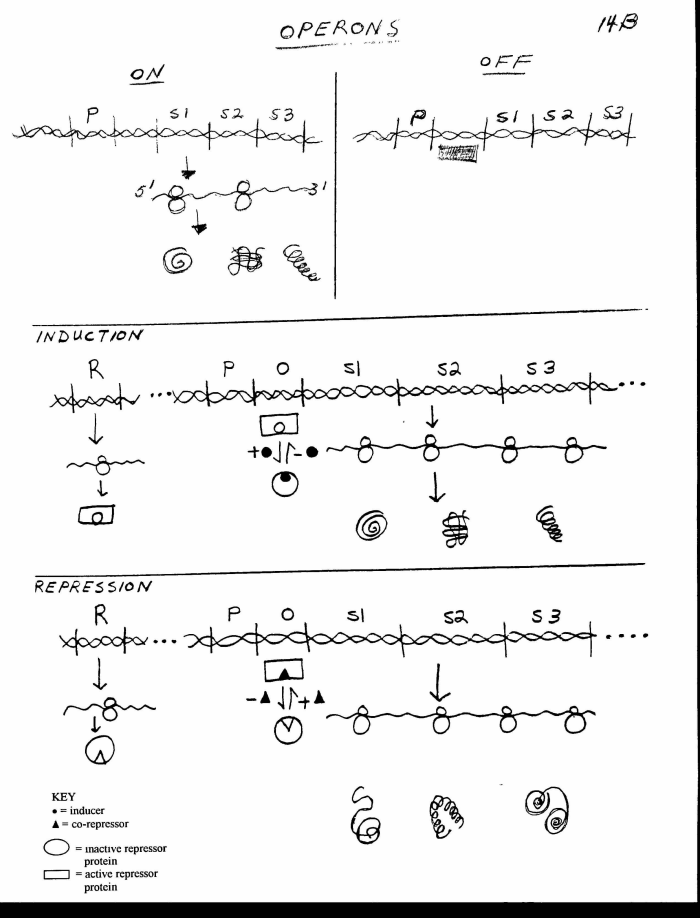
IV. Mechanism of Prokaryote Regulation (See handout 15B) -- Operons
Note: This mechanism was largely figured out by analyzing mutants. How it was done is fascinating, but complex, so we will explain the mechanism first, and then you can try your hand at predicting the effects of mutations. See E below for more details, and problem set 12 for examples.
A. How is co-ordinate control achieved? Upper Left Panel on handout -- idea of cluster or operon. (See Sadava fig. 16.10 (13.18) or Becker fig. 23-3.)
1. Genes regulated together are linked -- genes to be controlled co-ordinately (turned on and off together) are next to each other on the DNA.
2. Polycistronic mRNA. The linked genes are transcribed as a unit to give one single mRNA . One mRNA is made per operon (not one mRNA per gene), because all the genes in a cluster share a single promoter. An mRNA able to code for several peptides (mRNA that comes from several genes) is called polycistronic mRNA. (cistron = another term for gene).
3. Transcriptional Control -- Regulation is at level of transcription. The level of translation is controlled by regulating the synthesis of mRNA. This is the usual method for regulation of protein synthesis in prokaryotes.
Since mRNA has a short half life in prokaryotes, regulating mRNA synthesis controls the steady state level of mRNA. Translation per se (and degradation of mRNA) are not regulated here. (In some prok. cases and many euk. cases, these are regulated too.)4. Definition of an operon = group of linked structural (enzyme coding) genes that share common regulatory sites and that are transcribed as a single unit.
Note: The linked regulatory sites are always considered part of the operon; the gene for the repressor protein is not always considered part of the operon. Whether the repressor gene is considered part of the operon or not is usually clear from context; the role of repressor is discussed further below.
5. Punctuation. Reminder: DNA replication, transcription, and translation, have different stop and start signals. DNA replication starts at origins, transcription starts at promoters, and translation begins at start codons (AUG). Origins vs. Promoters was covered before. What about promoters vs. start codons?
a. mRNA has UTR's. It has leaders (untranslated region on 5' end before first AUG or 5' UTR) & trailers (untranslated 3' end or 3' UTR).
b. Numbers: Number of transcription starts (Promoters) for a message is one; number of translation starts may be many (one per peptide) in prokaryotes.
c. Translation of a polycistronic mRNA starts at multiple start codons. A ribosome assembles at the first AUG and starts translation. After each peptide is completed, the ribosome may continue down the mRNA to the next start codon and start a new peptide chain. Alternatively, the ribosome may detach (and disassociate into subunits) when it comes to a stop codon. In that case a new ribosome forms at the next start codon and starts translation of the next peptide.
B. How transcription of cluster is turned off -- Upper Right Panel of 15B -- Role of Repressor & Operator -- operon that is "off" (See Becker fig. 23-4, top panel or Sadava fig. 16.11 (13.19) top panel.
1. Role of operator (O) = DNA site to act as part of on/off switch -- binds repressor (regulator) protein when repressor is in appropriate or active form (rectangle on handout).
2. Role of repressor = other half of on/off switch (with O). Repressor is a protein that binds to operator and prevents RNA polymerase from binding to DNA and transcribing the operon. (Purves fig. 13.15 in 7th ed).
a. There is a different repressor protein for each operon. Repressor binds to specific sequence of DNA found in its respective operator.
b. Synthesis of repressor protein is constitutive -- gene is always on. (State of repressor protein varies, not the amount; see below.)
c. Terminology. The terms 'repressor' and 'repressor protein' are used interchangeably. The term 'repressor' is used in both induction & repression because the job of the protein is to turn the operon off. However some prefer to use the term 'regulator protein' instead of 'repressor protein' when referring to induction.
Question: Does the gene for repressor protein have a promoter? an operator?
C. How induction (and repression occur) -- Role of Effectors
1. Repressor protein is allosteric (has two forms) -- one that sticks to the operator and blocks transcription (active form = rectangle on handout) and one that doesn't (inactive form = round on handout). See Becker Fig. 23-5.
2. Repressor binds effector (inducer or co-repressor). Each repressor/regulator protein is unique in that it binds the proper co-repressor or inducer (see below) as well as the proper operator.
3. Effector determines which form the repressor is in. The amount of repressor protein present doesn't change (see above); the form repressor is in does change. The small molecule effector (inducer or co-repressor) shifts the balance between the two forms thus shifting the equilibrium between free (inactive) and bound (active) repressor and turning the operon "on" or "off."
4. How does repressor get on or off the DNA? The picture on the handout shows that the repressor is either "on" the operator (in rectangle form) or "off" the operator, (in circle form). There are 2 basic models for how the repressor gets on or off the operator. They are described below, but none of the problems in this course require you to know the difference between the two.
FYI, for those who like the details, there are two models for how an effector works:a. There is an equilibrium between free and bound "sticky" (rectangle form) repressor -- "rectangular" molecules are spontaneously coming on and off. The effector binds to the free repressor (not the repressor bound to the DNA). Binding of repressor and effector shifts the equilibrium between free rectangles and circles, which in turn shifts the equilibrium between free and bound rectangles.
b. The effector binds to the repressor on the DNA, changes its conformation, and causes it to move onto or off the operator. (In this model the repressor is always bound to DNA, but it moves from a random spot -- where it has no effect -- to the operator or vice versa.)
Older versions of the notes explained model a, but current evidence favors model b.
D. An example of Induction-- (see middle panel of handout 15B or Becker fig. 23-4 or Sadava fig. 16.11 (13.19). For an animation try http://vcell.ndsu.nodak.edu/animations/lacOperon/index.htm. Additional animations are listed on the links page. (There are multiple animations on the web and on YouTube. If you find an animation that is especially useful, please tell Dr. M. ) See next time for an animation of repression.
What are the characteristics of an inducible Operon?
Effector molecule (inducer) that binds to repressor protein prevents repressor from binding to operator -- causes rectangle form to change to circle form and fall off the operator.
Effector (Inducer) shifts following equilibrium to right:
"Rectangle form" of rep. protein ("sticky" form that binds to O) ↔ "Circle form" (form that doesn't bind to O)
Empty form of repressor protein (without effector) sticks to operator.
To review regulation so far, try problem 12-0.
E. Constitutive Mutants
1. What happens if repressor protein is mutant and doesn't bind to DNA at all? Will operon be on? off? Inducible or constitutive?
2. What happens if operator is deleted? Is it the same as above?
See
problem 12-3, part A.3. How do you test out the properties of constitutive mutants?
a. Many experiments and problems involve having a cell with two copies of an operon. (See below.)
b. How is this possible? Bacteria are haploid* -- each bacterium normally has only one DNA molecule (chromosome) with one copy of each gene or operon.
c. Bacteria can acquire an extra copy of a gene or an operon; the extra copy is usually on a plasmid. Such cells are called partial diploids.*(How cells acquire plasmids will be discussed next time.)
d. What are plasmids?
(1). Plasmids are mini-chromosomes that have 'extra' genes. Each plasmid has an origin of replication, so plasmids are replicated and passed on. (Details next time.)
(2). The 'extra' genes on the plasmid can be totally new or they can be additional copies of the genes already in the cell.
(3). A bacterium with a plasmid can be a partial diploid* -- it can have two copies of a gene or two copies of a whole operon. One copy will be its normal place on the chromosome and the other copy will be on a plasmid.
e. What use are partial diploids? The two copies do not have to be exactly the same -- one can be normal and one mutant, or they can both be different mutants. For example, suppose a bacterium has two copies of the lactose operon. Suppose one copy is constitutive and the other is inducible, or suppose both are constitutive. What should happen when you put the two operons together? Will both be constitutive? Both inducible?
*Terminology: A cell with one copy of every gene is called a haploid; a cell with two copies of every gene is called a diploid. A cell that is basically haploid, but has two copies of a few genes is called a partial diploid.
4. Use of Mutants. Study of the properties of constitutive mutants was how induction/repression was figured out by Jacob and Monod, who received the Nobel prize in 1965 for their work. Now you can try it the other way -- you can use your knowledge of operon function to predict the properties of mutants, both singly and in combination. See Chap. 12 of the problem book.
To learn how to tell the types of constitutive mutants apart, see problems 12-4 & Becker table 23-2.
F. Strong & Weak Promoters -- all promoters are not the same.
1. All Promoters are similar in structure and function -- all P's have to able to bind RNA polymerase and serve as signals to start transcription.
2. P's can be strong or weak
a. Weak Promoter → little (or infrequent) RNA polymerase binding → low levels of transcription → low levels of corresponding protein.
b. Strong Promoter → lots of (or frequent) RNA polymerase binding → high levels of transcription → high levels of corresponding protein.
c. Why does strength of promoter matter? The strength of the promoter determines how much mRNA can be made. Actual amount of mRNA made at any time depends on both strength of promoter and extent of repression or induction.
3. Example of strong vs. weak Promoters: P of lac operon vs P of lac repressor gene
a. Promoter of lac operon is strong. P of lac operon = P for the structural genes; controls production of polycistronic mRNA → enzymes for metabolism of lactose. Since this P is strong, you make a lot of mRNA and a lot of the corresponding enzymes.
b. Promoter of lac repressor gene is weak. P of lac repressor = P for the R gene; controls production of mRNA for lac repressor → lac repressor protein. Since this P is weak, you make only a little of the mRNA, and relatively little of the repressor protein.
c. Why does this make sense? You need a lot of the metabolic enzymes (if you are growing on lactose as a carbon and energy source) but relatively few molecules (100 or so) of repressor protein.4. Note difference between Roles of O (operator) and P (promoter). P determines what the maximum level of transcription is; O (plus Repressor) determines what percent of maximum is actually reached.
a. O (by binding to repressor) determines to what extent transcription (& protein synthesis) is "on" -- is protein synthesis running at full throttle or is it only partially turned on (or completely off)? Each individual operon or cell is probably "off" or "on" at any one moment. However, in an entire bacterial culture, not all cells are necessarily on or off. At intermediate levels of inducer, some cells may have their operon turned on and some may not. In these cells, some of the repressor protein is in the "rectangle" or active form, and some is in the "circle" or inactive form. There is some variation from cell to cell, and there is a threshold value for the amount of active repressor required to keep the operon 'off.'
Note: one molecule of active repressor (in the "rectangle form") per cell is not enough to shut down one operon. There has to be more than one molecule of active repressor protein per operon to be sure the operator is always occupied with a repressor protein molecule.
b. P determines the maximum level of transcription = level per culture when all operons are "on" and running at full throttle = level per cell when culture is fully induced.
See
problem 12-3, and compare parts A & B.Next time: Review of operons -- repression vs. induction. Then, how is bacterial (& viral) DNA passed on asexually? How do bacteria & viruses have sex, and how are the results of bacterial and viral crosses analyzed? (There will be a handout on all the details.)
© Copyright 2010 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY.
{kind=link}