p. 24 Add
the following note, which was omitted accidentally from the previous
edition:
*Note: a residue is that part of a
monomer structure that remains after it is incorporated into a polymer, i.e.
most of it.
p. 30. Problem 4-14 part A: 'How many net molecules of ATP will be generated...'
p. 34. Problem 5-6. Add the following:
Hint for part F: In the triose phosphate isomerase
reaction (rxn 5) does the phosphate group move?
p. 36. Problem 5-11, part B. The third sentence reads:
'This time the
same amount of NADH2 is oxidized and the same amount of oxygen is
used up, but the concentration of ADP
and Pi does not change.' Cross out the indicted section and
substitute instead: 'but you do not measure the changes in ADP and Pi.' (In the revised version of
B-5, you have to decide what will happen to the concentrations of ADP and Pi .)
Questions B-1 to B-4 are the same. Replace Question B-5 with the
following:
B-5.
In this experiment, you would expect ATP to be generated from ADP (in the
buffer) (inside the vesicles) (both) (neither).
p. 46. Problem 6-16, part D. Set up is the same, but change choices of answers as
follows:
This
information indicates that the native nucleic acid of this virus is probably (a
single strand doubled back on itself) (single stranded ) (double stranded) (any
of these) AND the DNA is probably (circular) (linear) (either way).
p.50 and thereafter -- the proper spelling is promoter, not promotor.
p. 61. Problem 8-9 has been rephrased to make it clearer. Here is the
revised version:
8-9. Suppose nondisjunction (ND) occurs during meiosis in a human
male. Assume that ND occurs only once per meiosis, at either 1st or
2nd division. Also assume that ND is equally likely at either
meiotic division, and that every meiosis produces 4 gametes (sperm). Now
consider the gametes that will end up with an abnormal number of sex
chromosomes. (Ignore the normal gametes.)
A. Which should be the most common type of gamete with an abnormal
number of sex chromosomes?
B.
Which should be the least common type of gamete with an abnormal number
of sex chromosomes?
The hint at the end is the same.
p. 91. Problem 11-11. Assume the bacteria have no plasmids.
p. 94. Problem 12-0, part C. In the list of possible answers, one choice has been
changed. Replace the choice 'binding of RNA polymerase to
promoter of greasy operon' with the choice: 'initiation of transcription from ........'
p. 102. Problem 12R-5, part D. The 2nd sentence should
read: The replication fork(s) should
move.....
p. 106. Problem 13-8, part B. You cut up your DNA
& run gels as in A, not
as on A.
In part B-2,
the last enzyme treatment should be EatIII
+ Eat IV, not EatII + EatIV.
Corrections to Answers
p. 146, Ans. to 2-20, part C. The answer has been changed to match the new sequences.
The new answer reads as follows:
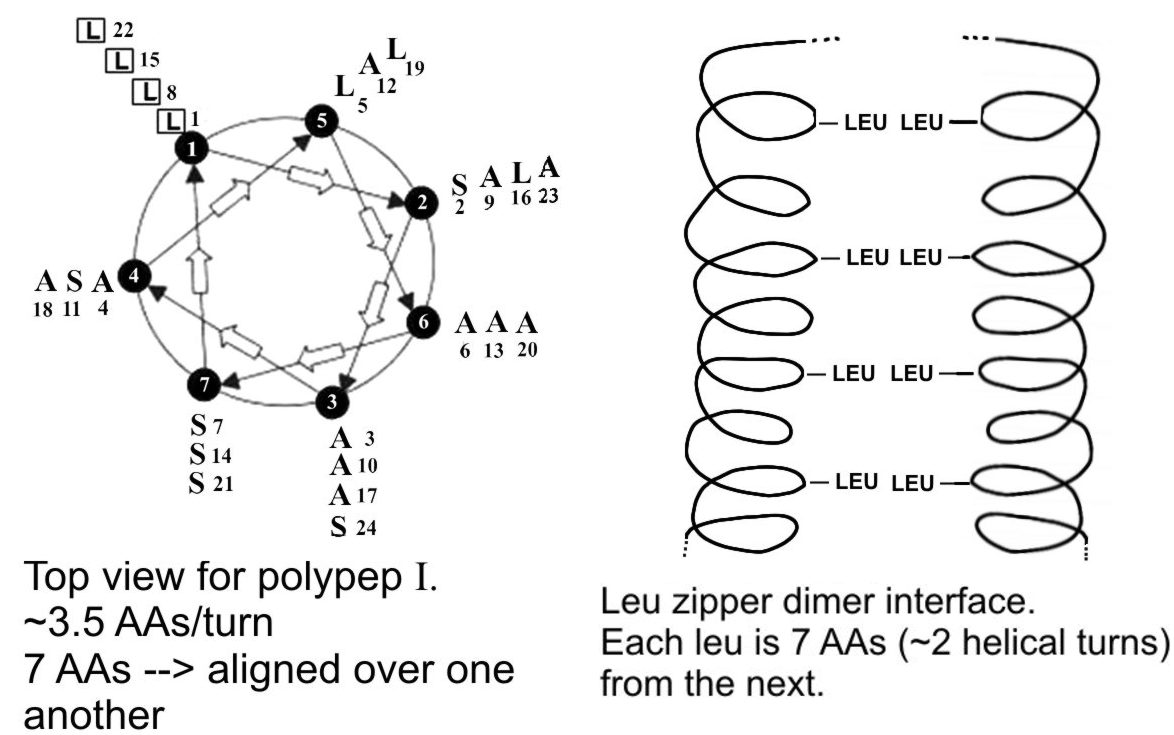
C. Both oligopeptides could form an alpha helical
secondary structure. The amino acid side chains protrude from the alpha helix.
In the case of polypeptide 4, there are 4 leucine residues that are spaced 7
apart (1, 8, 15, 22). Since there are ~3.5 AAs per turn, then these leucine
residues, being 7 AAs apart, would all be sticking out along the same side of
the helix, aligned one over (or beside) the other. (See left diagram below.) If the side of the alpha helix with these
4 leucines faces the exterior of the protein, then it could interact by
hydrophobic forces with the same face presented by an identical subunit of the
protein, thus contributing to the association of two subunits in forming the
homodimer. (See right diagram below.)
In oligopeptide II the leucine side groups cannot help connect subunits,
because the side chains would be pointing in various directions out from the
alpha helix. Serines (which could interact via hydrogen bonding) cannot help
connect subunits in either case, because the serines are distributed on all
sides in both oligopeptides.
Picture for Problem 2-20, part C.
p. 148. Ans. to new 3-6 & 3-7 are as follows:
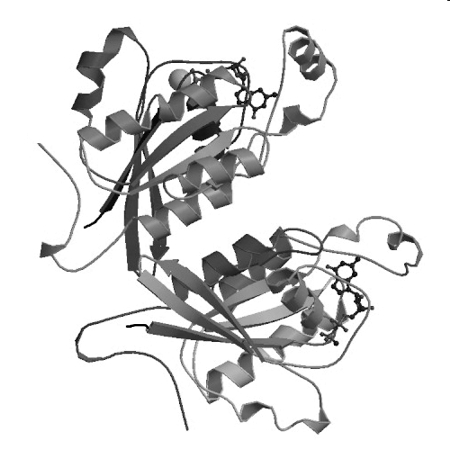
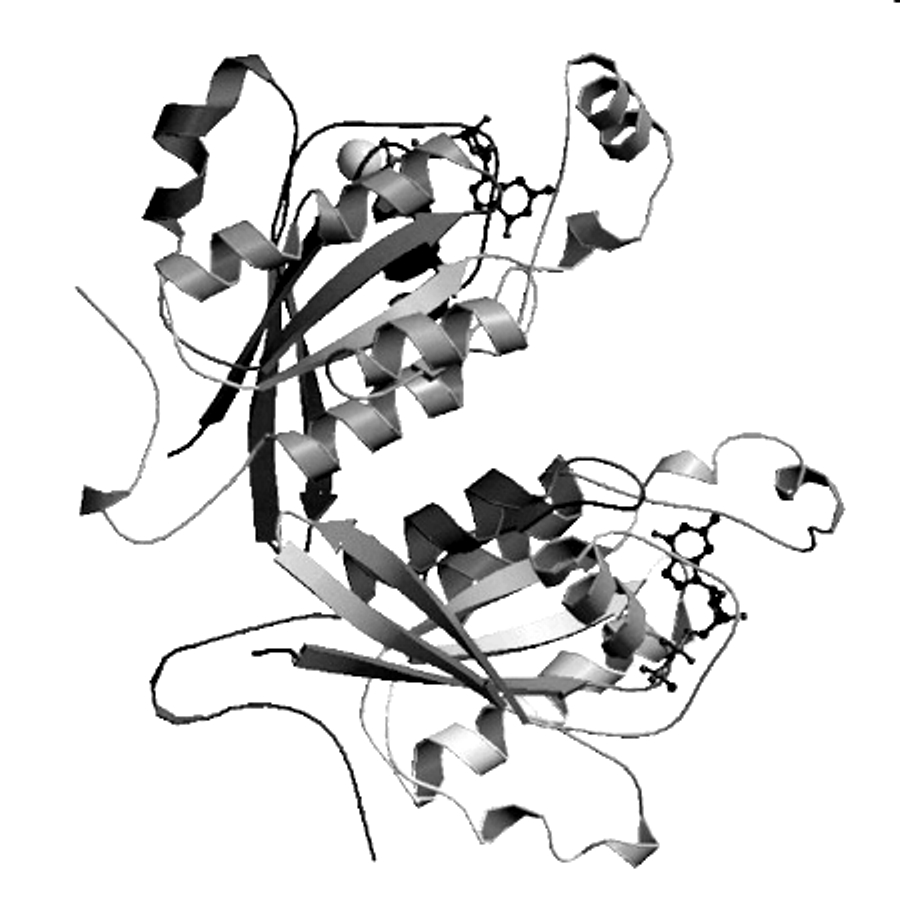
3-6. A & B. Two
polypeptides &. two subunits. (Each peptide = one subunit.)
C & D. 10 Alpha helices, 1 strand per helix.
E & F. Two beta sheets, with 10 strands total.
Each beta sheet is curved, so you might conclude that each curved sheet should
be considered two flat sheets at slight angles to each other. (In that case,
each subunit would contain one sheet with 3 strands and a second sheet with 2
strands, instead of one sheet with 5 strands.) That the 'two' beta sheets
really form one curved structure is shown most clearly in the bottom subunit.
G, H, & I. Two each. Each peptide chain has a carboxyl end and an amino
end, and binds one ligand molecule.
J. Both. Some strands are parallel and some are antiparallel. The parallel ones
are seen most clearly in the top subunit.
3-7. A. A straight line, since if S << Km, Vo will
be proportional to [S]. At very low [S], Vo ≈ k3 Eo [S]/Km
B. A straight (horizontal) line, since if S>>Km, Vo
will be constant. At very large [S], Vo ≈ k3 Eo.
C. 10,000, since Kd
~ Km and Ka = 1/Kd . (Kd = Keq for
disassociation of ES or the reaction ES ↔ E + S; Ka
= Keq for the reverse reaction.)
D. Higher. Remember we are
asking about how the real Ka relates to the Ka calculated
using the Km , not how Kd relates to Km. Kd
can be lower than Km , so the real Ka or 1/Kd
can be higher than 1/Km. 'The reasoning is as follows: Km
= (k2+k3)/k1 = k2/k1 + k3/k1 = Kd+k3/k1. Therefore Kd = Km-(k3/k1),
and Kd can only be lower than Km (usually it is
insignificantly lower). Since Ka = 1/Kd , if Kd
is lower than than expected, then Ka can only be higher than
expected (higher > 1/Km).
E-1. Stay the same. The Vmax/2
has changed, but the amount of S it takes to get there is the same.
E-2. Non-competitive (because Km stays the same, but the Vmax
is affected).
E-3. No for this problem; yes for problem 1. A competitive inhibitor (as in problem
1) should look like the substrate; a non-competitive inhibitor (as in this
problem) does not bind to the substrate binding site and should look different
from the substrate.
p. 154, Ans to Problem 4-7,
part B. Add the following::
Note
that starting concentrations of K and M are as in part A, not as described at the start of the problem.
p. 157, Ans to Problem 4-14, part A. Add: You put in one ATP to
convert glycerol to glyceraldehyde, but you get two ATP back per glyceraldehyde
phosphate broken down.
p. 158, Answer to Problem 4-18, part A (i). Add:
The radioactive
acetate combines with OAA to give citric acid (citrate) before any CO2
is lost or other radioactive compounds are formed.
p. 160, Answer to Problem 5-6, part F. Add:
Note that in reaction 5, the conversion of DHAP to glyceraldehyde P, the P doesn't move, so you have to turn the molecule upside down.
p. 162, Answer to Problem 5-11, parts B-1 & B-5 are redone.
(The other answers are unchanged.) Here are the revised answers to B-1 &
B-5:
B-1.Electron transport is
occurring (and a proton gradient is being generated; see below.) There is not
enough info given to indicate if ox. phos. is occurring or not, but it should
be okay if all the components in the membrane are working. If the membranes are
not damaged, ATP can be made with the membranes in either orientation (if the
appropriate substrates are on the appropriate sides). For an example, see
problem 5-16, and also B-5 below.
In normal undamaged membranes, ox. phos. and electron transport are
coupled; if you block one, the other backs up. Since electron transport is
normal, we assume that ox phos is occurring too, and everything is working as
it should. (Why does blocking ox. phos. block electron transport? If ox. phos.
does not occur, the H+ gradient usually builds up to very high
values and electron transport is inhibited as a result. This has not happened
here.)
If the membranes are
damaged, as often happens in real experiments, then ox. phos. and electron
transport can get uncoupled. Protons can leak out, the H+ gradient
doesn't get too high, and electron transport can continue without ox. phos.
B-5. In the buffer, assuming the membranes are not damaged. Remember the
membrane is reversed, relative to part A and to the usual orientation. Since
the topology is reversed in this experiment (relative to the normal), the H+
are pumped into the vesicles, and
flow out, through ATP synthetase,
driving phosphorylation of ADP to ATP.
If the membranes are damaged, then electron transport and ox. phos. can
be uncoupled, as explained above. Even though electron transport and H+
pumping continue, H+ can leak back out, bypassing ATP synthetase,
and no ATP will be made on either side.
p. 172, Answer to problem 6-16, part D. The
revised answer is as follows:
D.
Double stranded & linear.
(Double stranded RNA is not found in normal cells, but is found in some viruses
and infected cells.) The heat treatment breaks hydrogen bonds, not covalent
bonds. Since the molecular weight is reduced to 1/2 by heat, the native
molecule must consist of 2 complementary strands connected by H bonds that can separate completely when
heated. When the nucleic acid is cooled down, the complementary strands
match up and the H bonds are reformed. If the DNA were circular, the two strands would be wound around each
other in a double helix, and would not separate when the DNA was denatured.
(You would have two linked rings.)
p.
174, Answer to problem 7-7, part C. Replace 'One way' with 'One way to do the experiment in A.'
p. 175, Answer to problem 7-14. Replace 'lys-tRNA' and 'tRNA-lys' with
'tRNAlys.'
Answer to Part A: The
last sentence of part A should read: 'Only two tRNAs are bound to the
ribosome at any one time (either in A & P or in P & E), but you need a
third tRNAlys because the E site will not empty until a lys-tRNAlys
arrives in the A site.'
p. 198, Answer to problem 10-5, parts C
& D. The answers have been rewritten slightly to make them clearer. The revised answers are as follows:
C.
You need to calculate the fraction that are triple homozygotes (gg bb dd) from
a cross of two triple heterogyzgotes (GgBbDd X the same). Simplest way is to
consider each gene separately.
Answer
= (fraction of offspring that are homozygous for 1st gene) X
(fraction homozygous for 2nd gene) X (fraction homozygous for the 3rd
gene) = fraction that are gg X fraction bb X fraction dd = 1/4 X 1/4 X 1/4
=1/64.
D.
Ans. = (fraction that are G_) X (fraction that are Bb) X (fraction that are dd)
= 3/4 X 1/2 X 1/4 = 3/32
p. 198, Answer to problem 10-8, part B. Add the following:
Note that 'linked' does not
mean 100% linkage (no crossing over & no recombinants). It simply means
there are more parentals than recombinants -- no independent assortment. See
explanation to 10-10, A. (2).
p. 202, Answer to problem 10-19, part A-1. Add
the words in blue. 'If they look the same, the plants have the same phenotype. Plants with
the dominant phenotype can have different genotypes (heterozygous -- in F1, or homozygous -- in Parents).'
p. 207. Answer to
problem 11-8, part B. Part ii
should read:
ii) 4% = both normals &
double mutants. The parentals were single mutants, with mutation 1, 2, or 3.
Each crossing over event between 2 single mutants produced a double mutant and
a 'mutant-less' normal.
The explanation to part iii
had a serious typo in it. (The term in parentheses in line 4 should be
'mutant', not 'non-mutant.' ) Here
is a corrected version: :
How to decide between
complementation and recombination in part iii? If function is restored without
any opportunity for crossing over, you know it was complementation. Otherwise,
it is sometimes difficult to tell -- you have to look closely at the details in
the question to see how to rule out one or the other in any particular case.
(There is no quick foolproof method.) In this problem, most of the progeny are
mutant and only a few are recombinant. This makes sense if complementation
allowed infection to proceed to the point where some crossing over could occur.
Only a few progeny DNA molecules engaged in crossing over, so the rest are mutants,
produced as a result of complementation.
p. 209, Answer to problem 11-14, part B. The arrowheads in the diagram are not
all properly aligned. However the rest of the diagram is correct.
p. 214, Answers to problem 12-0, part C.
For item (2): Initiation of RNA synthesis by RNA
polymerase will increase, but RNA polymerase binding may not. RNA
polymerase may already be bound, but if it is, it can't do anything
because initiation of RNA synthesis is blocked by repressor.
{kind=link}
{kind=link}