Lec. 8. Biol C2005/F2401 2004 L. Chasin Oct. 3, 2006
Last updated:
Monday, October 09, 2006 01:35 PM, with some minor
corrections
© Copyright 2006 Lawrence Chasin and Deborah Mowshowitz Department
of Biological Sciences Columbia University New York, NY
Gycolysis cont.
Anaerobiosis
Fermentation
Energy yield
Krebs Cycle (TCA cycle, citric acid cycle)
FAD
electron transport chain
mitochondria
chemiosmotic theory
proton pump
oxidative phosphorylation
substrate level phosphorylation
one handout
(Glycolysis)
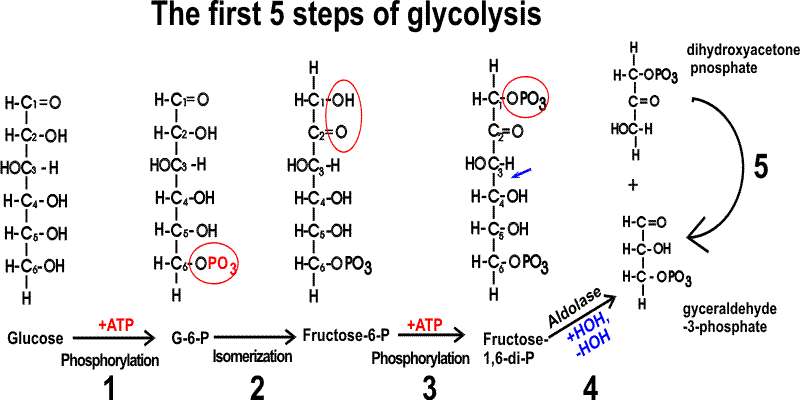
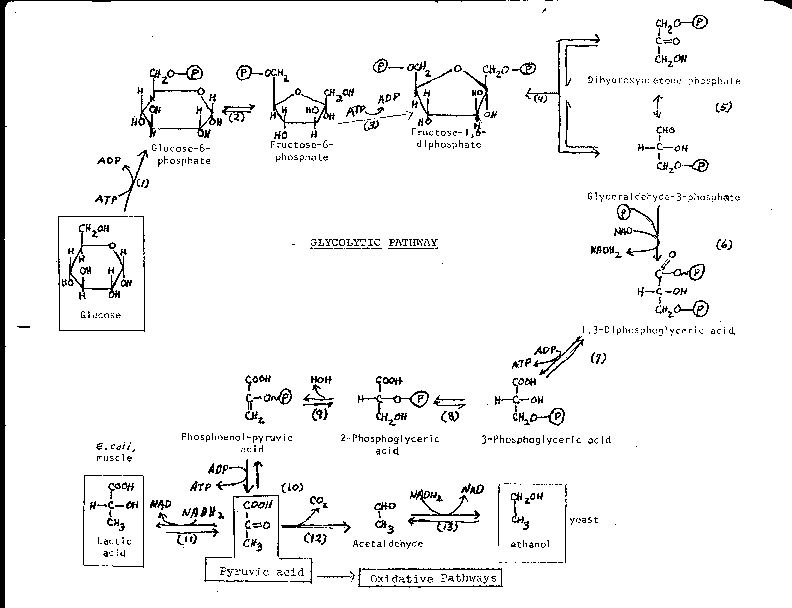
GLYCOLYSIS:
I'll show the first few reactions with the
sugar in open chain form, because I think it is easier to see what's going on.
1. a phosphorylation (kinase enzyme)
2. an isomerization
3. phosphorylation again... note that we have now used 2 ATPs: rather than generating energy, we are consuming it, so far.
4. hydrolysis (For more about this reaction see Problem 4-16)
5. isomerization of dihydroxyacetone phosphate (as we saw in step 2, but in reverse, transforming the ketone into the aldehyde; and swing it upside down when done: you have another molecule of glyceraldehyde-3-phosphate): note now we have 2 of everything for each glucose molecule that entered the pathway, this will be true from this point on.
So, so far, through rxn 5, we've produced 2 molecules of glyceraldehyde-3-phosphate (GAL-3-P) for each molecule of glucose we used. AND it so far has COST us 2 ATP's This ATP debt is a loose end that we will have to deal with sooner or later.....
6. Step 6 is an oxidation, GAL-3P plus another phosphate to form 1,3 diphosphoglyceric acid, (or 1,3-diPGA). {Q&A}
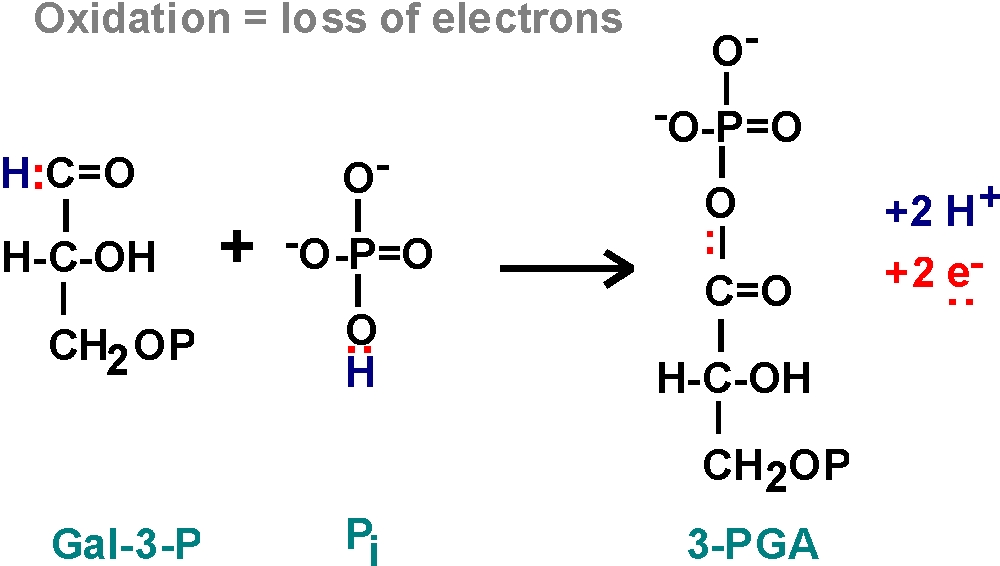
OXIDATION, the loss of ELECTRONS. We saw it before in the formation of the disulfide bond - there the loss of two electrons was in the two hydrogen atoms that were taken away. Here, in reaction 6, we have 2 electrons to be lost from the reactants, GAL-3-P and phosphate:
(NAD)
Two electrons have been lost. The protons are not important in oxidation,
they are sometimes there with the electrons, and sometimes not.
{Q&A} These electrons must go somewhere. They are taken up by the
OXIDIZING AGENT which itself will
get reduced. As you can see from the
NAD
handout, this is a compound called NAD, or
NICOTINAMIDE ADENINE DINUCLEOTIDE
[Purves picture]:
The nicotinamide part is also the vitamin niacin. Niacin sounds less ominous.
In getting reduced, NAD can accept two electrons, but only one proton. The other proton goes into solution as a hydrogen ion. But it is the electrons that are important in oxidations.
Instead of w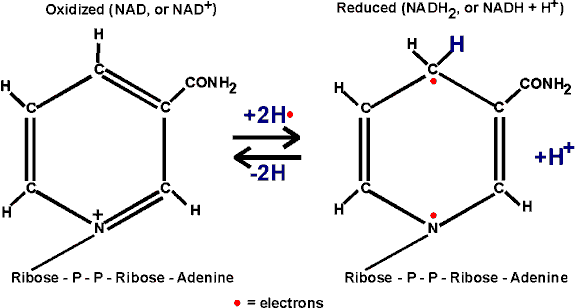 riting the reduction of NAD as NAD+
--> NADH + H+, we will simply write: NAD --> NADH2, referring
to the two H's that came from glyceraldehyde-3-phosphate, despite the fact that
both protons did not end up on NAD.
{Q&A}
{Q&A}
riting the reduction of NAD as NAD+
--> NADH + H+, we will simply write: NAD --> NADH2, referring
to the two H's that came from glyceraldehyde-3-phosphate, despite the fact that
both protons did not end up on NAD.
{Q&A}
{Q&A}
This rxn. 6 is rather complicated, involving a phosphorylation as well as an oxidation. The phosphorylation did not require an ATP, but it did require an NAD.... so now have a new loose end analogous to the ATP/ADP situation, we have to worry about restoring NADs from NADH2's as well as ATP's from ADP's.
But at least we have something of value here, a high energy phosphate, as you can see the squiggle at the top of 1,3-di-PGA on your glycolysis handout. That means that enough energy can be released from the hydrolysis of this phosphate even to drive the phosphorylation of ADP in a coupled reaction.
Let's take the money and run... this high energy bond is cashed in the next reaction, rxn 7. Note that the top carbon is now a carboxylic acid in 3-PGA, whereas it had been an aldehyde in GAL-3-P. This change is the result of the oxidation that took place in reaction 6. {Q&A}
So we've now paid off one of the 2 ATPs the
debt we ran up at the start .….
Wait a minute, actually, we're all paid up (since we have two di-PGA's
for every mole of glucose that started down the glycolytic pathway).
Continuing along:
8. Isomerization of 3-PGA to 2-PGA.
9. Dehydration, water removed. The result is an unstable compound, phospho-enol pyruvic acid (PEP), one whose hydrolysis can result in the release of a large packet of free energy.
[2-PGA, as well as PEP, is at a higher energy level than pyruvate, but the shift in atoms allows cash-in of ATP along the way, since the phosphate bond is a high energy bond in PEP.]
10. Transfer of the phosphate to ADP (X2), resulting in PYRUVIC ACID (pyruvate), which can be considered the end point of glycolysis. [Purves picture], [another Purves picture], [yet another Purves picture]
So after the 10 steps of glycolysis we have 4 ATPs produced and 2 invested, for a net gain of 2 ATPs produced from ADP. So glycolysis does produce energy in the form of ATP. And the overall reaction will run far to the right, as the Delta Go from glucose to 2 pyruvates is -18 kcal/mole (even taking coupled reactions producing the 2 net ATP into account). That is,
1 glucose + 2 ADP + 2 Pi + 2 NAD <--> 2 pyruvate + 2 ATP + 2 NADH2
Delta Go = -18 kcal/mole
The diagram below shows Delta Go's for the individual steps in glycolysis.
(For a different way of looking at it, see the chart of Delta G's drawn by Purves [Purves picture])
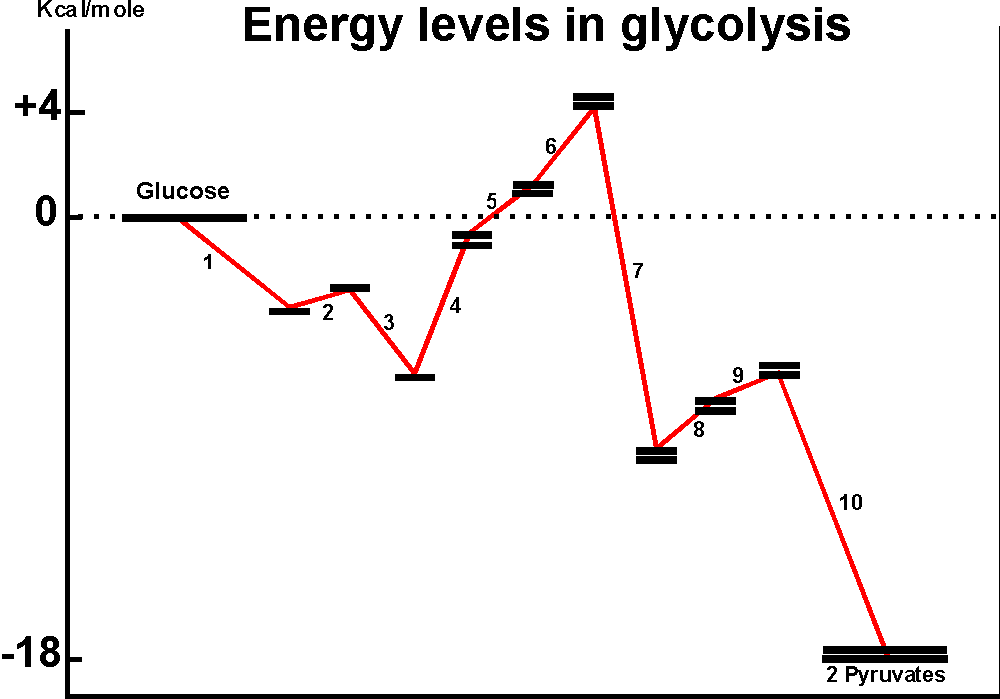
Although the pathway overall is quite favorable, some individual reactions are quite unfavorable, the most extreme being reaction #4. The energy-requiring reactions in this case are being pushed by the build-up of reactants by the more favorable reactions before them, and they are being pulled by the withdrawal of products by the more exergonic reactions further downstream. The actual Delta G's will be influenced by the drain of products, so that the second term in the equation for Delta G is being brought into play here. {Q&A}
Delta G = Delta Go + RTln([products]/[reactants]), where RT~0.6 (in kcal/mole)
For instance, if the products are drained such that the ratio of P/R reaches down to 0.00001, this produces ~ 7 kcal/mole of negative Delta G , enough to balance out the unfavorable Delta Go of +6.8 for rxn. 4.
This indirect affect on the Delta G is the second method the cell utilizes to carry out an individual reaction that has an unfavorable Delta Go.
So we have 2 methods:
1.) DIRECT COUPLING of the unfavorable reaction to an energetically favorable one to produce a new coupled reaction with a net negative (favorable) Delta Go (as in the hexokinase reaction #1).
2.) INDIRECTLY, via the WITHDRAWAL OF PRODUCTS or buildup of substrates {Q&A}
SO, we have our overall negative Delta Go , and we have generated net ATP, so we should feel pretty good, except for one thing: we have an important loose end to tie up. We borrowed an NAD to oxidize Gal-3-PO4. That was a key reaction in the path, as oxidations are usually accompanied by large changes in free energy. We were able to get a PO4 added to our 3-carbon compound, and this phosphate was the one that was used to phosphorylate ADP in the very next step (#7). We used NAD, and it became reduced to NADH2. So now we must consider how we are going to repay that debt. Otherwise the small stores of NAD in the cell would very soon all be converted to NADH2 and glycolysis would quickly grind to a halt.
We need an oxidizing agent to oxidize NADH2. A great one abounds: O2.
It readily takes up electrons, for instance from Fe++ to make it Fe+++ as steel goes to rust.
(Anaerobiosis)
Does E. coli have access to oxygen? In the
lab, yes. We vigorously and constantly shake the E. coli cultures on mechanical
shakers to get air dissolved in the culture medium to provide a constant source
of oxygen, called an AEROBIC state. In the gut, sometimes yes, sometimes no
(crowded). Do our own cells have access to oxygen? Sure, through the lungs via
the blood vessels to all tissues. But when you are running across campus to
class so as not to miss the first golden words here, your muscles may need ATP
faster than you can deliver oxygen to them for NADH2
oxidation, your muscles will be in an AN-AEROBIC
state. And are many organisms that live in naturally anaerobic environments, in
mud at the bottom of rivers, e.g. So let's first consider the anaerobic case
when no oxygen is available for the oxidation of NADH2. Under
these circumstances the cell must make do with what it has available, which is
mainly: 2 pyruvates. Fortunately, pyruvate itself is able to act as an oxidizing
agent, as seen in rxn #11, where it accepts electrons into its C=O bond, adding
two hydrogens as well, to form lactate, or LACTIC ACID.
It gets the electrons from our NADH2 molecule, which is really NADH and H+, so
it is recapturing one proton (an H+ ion) from the aqueous pool. Even the
Delta Go is favorable, and we get our NAD regenerated from NADH2. The NAD
shuttles back and forth then, getting reduced in rxn 6 and reoxidized in rxn 11
over and over and over again, while glucose runs down to lactate, and ADP's are
converted to ATPs to power cell division for E. coli or running up stairs for
humans even in the case of insufficient oxygen.
It is thought that it's the build-up of acidity from lactic acid that makes your muscles hurt if they are doing anaerobic glycolysis too long (acidity sensed as pain by neurons in the muscle).
(Fermentation)
So pyruvate is a crossroads: If no oxygen,
then if you are E. coli or humans, you carry out a lactic acid
fermentation...[Purves
picture], [another Purves picture]
If you are yeast, there is a variation on this theme, you break down pyruvate first to acetaldehyde and CO2, which is not an oxidation, but then use the acetaldehyde as an oxidizing agent for NADH2, with the product here being the 2 carbon alcohol, ethanol (rxns 12 and 13). Beer drinkers appreciate this variation, as lactic acid beer would be pretty bad, and probably not even produce those psycho-pharmacological effects for which ethanol is famous. {Q&A}
Just as in the case of ATP, the NAD - NADH2 case is one of re-generation, not generation. Once a little NAD is made, it can shuttle back and forth millions of times getting alternately reduced and oxidized. [
Let's consider for a change an alternative source of carbon and energy for E. coli, glycerol. Remember glycerol is the tri-alcohol that serves as the backbone for triglycerides (fat) and phospholipids.
See glycolysis handout. On PowerPoint graphic, glycerol is metabolized as follows:
glycerol + ATP --> glycerol-1-P
glycerol-1-P + NAD --> DHAP (= dihydroxyacetone phosphate) + NADH2
DHAP --> continues in glycolysis (rxn 5). See recitation problems #3, problem 2.
Under aerobic conditions, no problem, the NADH2's produced will get re-oxidized by oxygen, as we will see. And so E. coli grow just fine in a glycerol minimal medium using glycerol instead of glucose.
How about under ANAEROBIC conditions?
We used two NADs to get down to pyruvate from glycerol, but we get back only one in going to lactate. If we try to run the lactate fermentation using glycerol as our only carbon and energy source, we will grind to a halt as all our NAD ends up as NADH2. (You will reach a state where all the NAD is in the form of NADH2, and there is no pyruvate left, just DHAP waiting for an NAD that is not there.) So, although E. coli will grow just fine on glycerol in the presence of air (oxygen), it will NOT, in fact, grow on glycerol in the absence of air (anaerobically). So: these loose-end debts are real, there is no magic, we must play by the rules.
See Problem 4-15 - why was he fired?]
Consider the efficiency of glucose fermentation:
For: glucose--> 2 lactates,
without considering the couplings for the formation
of ATP's (no energy harnessing):
Delta Go = -45 kcal/mole.
{Q&A}.
Out of this comes 2 ATPs, worth 14 kcal/mol. So the efficiency is about 14/45 = 30%, which is not bad, about what a gasoline engine can do.
Where did the other 31 kcal get to? They are released as heat, which is why, after you've run fast to class, in addition to the lactic acid PAIN in your legs, you are also SWEATING.
Now taking the ATPs into account, the overall Delta Go is about -45 + 14 = -31 kcal/mole, so the lactic acid fermentation runs essentially completely to the right (or clockwise) as written.
(Energy yield)
The efficiency is pretty good, but on the
other hand the YIELD is poor. What do I mean
by YIELD? Well, glucose has a lot more chemical energy in it than we are tapping
here. For example, if we BURN glucose (i.e., react it with oxygen) and measure
the calories of heat given off, we get no less than 686 kcal/mole. Compare that
to the measly 45 kcal/mole we got from converting glucose to two lactates. The
lactic acid that we are throwing away at the end could be used for more energy,
but in the absence of oxygen there is no way to use it.
With oxygen, it's another story .............and our next chapter.
[To review glycolysis and fermentation, try Problems 4-11 and then 4-13.]
(Krebs Cycle (TCA
cycle, citric acid cycle) )
So now let us turn to the case when
oxygen is present. In this case we are
headed to the complete oxidation of glucose to 6 molecules of CO2 and
6 molecules of water:
C6H12O6 + 6 O2 ---> 6 CO2 + 6 H2O (write high on board and do not erase)
The Delta Go for this reaction is -686 kcal/mole, and we are going to hope to get a lot more ADP --> ATP conversions out of this.
In the presence of oxygen, glycolysis down to pyruvate is the same. But the fate of pyruvate is now different.
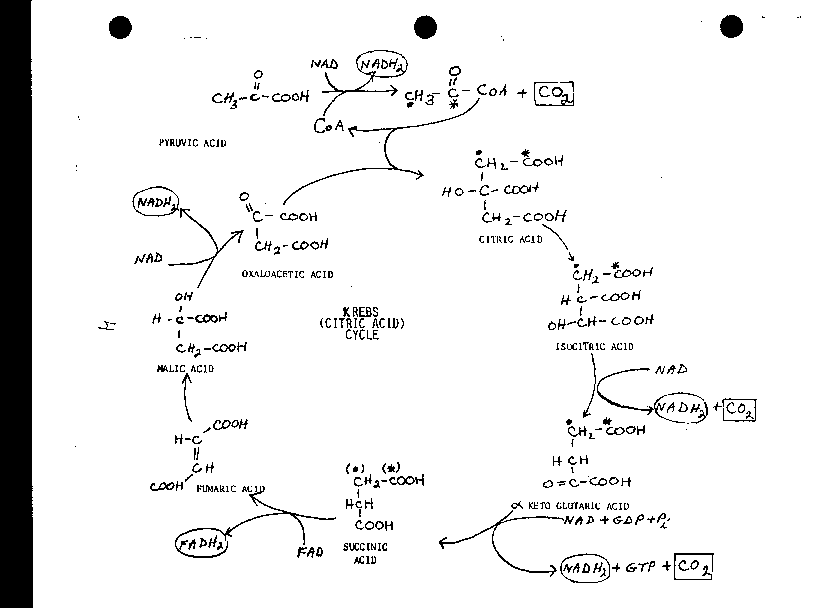
THE KREBS CYCLE
Rather than heading toward lactate or ethanol, as shown at the bottom of the glycolysis handout, the pyruvate jumps to another handout: the KREBS CYCLE. Keep in mind that by forgoing the reduction of pyruvate, we have not satisfied our loose end of NADH2 accumulation from the oxidation step (step 6) in glycolysis.
The fate of pyruvate is now different, it will enter a series of reactions known as the Krebs Cycle, (also TCA cycle, or citric acid cycle) in which all of its carbons will indeed end up as CO2. However, as we are about to see, its H's will not be converted to water here, and very little ATP will be produced here. [Purves picture], [another Purves picture]
Please follow along in the Krebs Cycle diagram as the reactions are discussed. At the top of the Krebs Cycle diagram, we see our pyruvate entering from the top left, in a reaction that is not part of the cycle of reactions seen below (it is sometimes called the entrance reaction). Before entering the cycle, pyruvate undergoes a relatively complicated reaction involving an oxidation, once again using NAD as the oxidizing agent, as well as a DECARBOXYLATION, the splitting off of the carboxyl carbon as CO2 (as when ethanol was made in yeast) and leaving a 2-carbon acetate group. So here is some CO2 produced, which is what we expected from the oxidation of glucose. In addition, a new co-factor makes an appearance: Coenzyme A (abbreviated CoA), a sulfur-containing small molecule [pADP-pantothenate-SH] which becomes bound to the acetate in a thioester linkage. A thioester is analogous to an ester except a sulfhydryl is one of the reactants instead of an alcohol, so a sulfur atom takes the place of an oxygen. A thioester contains a high energy bond, and so should be a SQUIGGLE (see Becker 6th Ed. Fig. 10-7 for the exact structures, if you wish). It is acetyl-CoA, the product of this dehydrogenation of pyruvate, that is the compound that now enters the Krebs Cycle proper. [Purves 7.9]
Acetyl-CoA adds to a molecule of oxaloacetate, a 4-carbon keto-dicarboxylic acid (across its carbonyl), to produce citrate, a 6-carbon tricarboxylic acid (thus the name tricarboxylic acid cycle or TCA cycle as a synonym for the Krebs Cycle). CoA is split off in the course of this reaction. The high energy bond to CoA is utilized to help drive this otherwise endergonic synthesis of a 6C molecule. The free CoA is regenerated and so is not consumed.
Now we have a new loose end, however: OA (oxaloacetate). We have introduced, borrowed, this molecule, much as we borrowed an NAD, so it's now OA plus 2 NADs; no, really 4 NADs and 2 OA's (per glucose molecule). We must pay back these debts by the end of our path (which remember now is going to be glucose going to CO2).
We have labeled the carbons of acetyl-CoA with an asterisk and a dot so that we can follow them as they go through this set of reactions. In the laboratory, it is also possible to use organic molecules labeled in this way, by using molecules in which particular carbons or hydrogens have been replaced with their radioactive counterparts, or isotopes, for instance 14C instead of the usual 12C or 3H instead of 1H. Radioactivity counters can then be used to track and measure the appearance of the isotopes in various purified intermediate compounds.
The citrate is isomerized to isocitrate; note the movement of the hydroxyl group from the middle to one end. The isocitrate is oxidized once again using NAD and it's simultaneously decarboxylated to produce alpha-ketoglutarate and now our second molecule of CO2. We need to get 3 from our 3-carbon pyruvate molecule. So this is 2. Except the actual carbon is not from our acetyl-CoA, i.e., from our pyruvate, but rather from the OA that we borrowed. Looks like it may be a bad debt: here we've borrowed an OA and now we've blown it into CO2.......How are we ever going to pay it back? Oh well, let's go on.
Next we again have an oxidative decarboxylation, from the 5-carbon alpha-ketoglutarate to the 4-carbon succinate. This is actually a set of two reactions, as can be seen in the Becker text. Once again the CO2 comes not from the acetyl-CoA carbons, but from a carbon atom originally in OA. But here we have a new and welcome feature, the production of GTP from GDP. The free energy from this oxidation is coupled to the phosphorylation of GDP with Pi. GDP is a compound, a nucleotide, related to ADP, the structure being the same except for the substitution of the guanine ring for the adenine ring. The production of GTP is energetically equivalent to producing ATP, since:
GTP + ADP --> ATP + GDP, Delta Go = 0
So we finally get some ATP here, 2 moles per mole of glucose, equal to what we netted in glycolysis. {Q&A}
The mechanism of this coupling is less obvious than those we saw in glycolysis (where the substrates were phosphorylated). Here the inorganic phosphate and GDP are both bound by the enzyme as part of the overall reaction.
[not responsible for this
mechanistic detail:
aKG + NAD + CoA --> CO2 + NADH2 + Succ~CoA;
Succ~CoA + Pi + E --> CoA + E-succ~P --> E~P + Succinate,
E~P + GDP --> E + GTP ]
(FAD)
Next the succinate is dehydrogenated across its central 2 C's, producing fumaric
acid This is the reaction we discussed earlier illustrating enzyme
specificity. Here the oxidizing agent is FLAVIN ADENINE
DINUCLEOTIDE (FAD), rather than NAD.
FAD is a better oxidizing agent (it is more easily reduced) than NAD; the Delta
Go for this reaction using NAD would be highly unfavorable,
whereas with FAD it is much more favorable, about a 10 kcal/mole
difference. So now we have to add 2 FADH2's to our debt
LIST. Our debt list now includes: NAD's, FAD's, and OA.
Continuing, we add water across fumarate's C=C double bond to get malate, and then once again dehydrogenate, i.e., oxidize, using NAD to get the 4-carbon dicarboxylic keto acid OA.
So we can pause here, our OA has been regenerated, and is ready to take on another acetyl-CoA. We have utilized one pyruvate and have released 1, 2, 3 CO2 molecules. We have carried out 5 oxidations per pyruvate, 4 with NAD and 1 with FAD, 4 oxidations in the cycle proper and one oxidation getting into the cycle. We produced one ATP equivalent per pyruvate. And we have accumulated 4 NADH2's and 1 FADH2 per pyruvate (double all these for a per glucose accounting). But we have paid back our OA debt, not with the original OA molecule, but no one will ever know the difference (except perhaps some nosy biochemist with radioactive isotopes).
| In Krebs Cycle |
per pyruvate |
per glucose |
| CO2 released |
3 |
6 |
| oxidations: |
5 |
10 |
| NADH2 produced |
4 |
8 |
| FADH2 produced |
1 |
2 |
| ATP produced (as GTP) |
1 |
2 |
| O2 consumed |
0 |
0 |
Our labeling showed that the CO2 molecules produced in this turn of the cycle were not derived from the acetyl-CoA. The acetyl carbons have ended up on either the top or the bottom of the OA that was regenerated. We don't know which end because fumarate is a symmetric molecule, and the water addition forming malate could have produced a hydroxyl on either the labeled or the unlabeled end.
So far we've gotten precious little energy out in the form of ATP, and we still have the NAD and FAD to pay back. And oxygen has not been involved yet. The electrons of glucose (entering here as pyruvate) have not been delivered to O2, but are still on the way-station of NADH2 and FADH2.
To review the Krebs Cycle, try problem 4-17 (4-12 in 16th ed.)
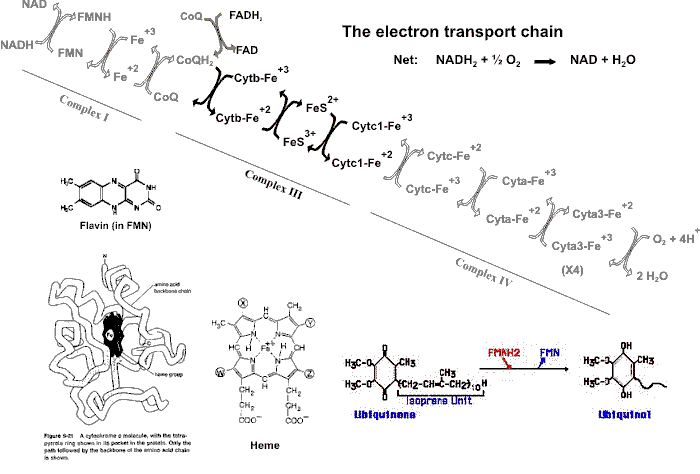
(electron transport chain)
In the next part of the story, NADH2
and FADH2 will pass their electrons on to oxygen AND we will
get a lot of ATP from this passage of electrons. This oxidation of NADH2
and FADH2 will return them for further action as NAD and FAD.
So let' s pass them on to oxygen:
NADH2 + 1/2 O2 --> NAD + H2O, Delta Go = -53 kcal/mole
53 kcal/mole released, TOO high. Too much energy released: if it were used in one fell swoop of the usual coupled reaction, we would get only a single ATP's worth, 7 kcal/mole, from this 53 kcal/mole, and we'd release a LOT of heat besides. It would be better if we could break up this 53 kcal/mole into smaller bits to use. The scheme for breaking up the free energy change involved in the reduction of oxygen involves passing the electrons from NADH2 and FADH2 not directly to oxygen but rather through a chain of intermediate transfer steps. This chain of steps is called the ELECTRON TRANSPORT CHAIN (E.T.C.).
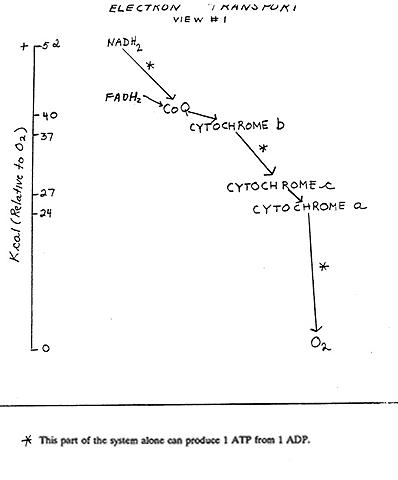
Let's look at View #2, on the ETC handout: The electrons from NADH2 are seen to be passed to various ELECTRON CARRIERS of the electron transport chain, in a precise sequence of transfers (here simplified, see Becker).
Some of the participants in this chain are:
- Proteins = Iron-sulfur protein (in which iron as Fe+++ (ferric) accepts the electrons).
- Coenzyme Q (CoQ) , a small molecule, hydrophobic, lipid soluble (also called ubiquinone).
- CYTOCHROMES: cytochromes b, an Fe/S protein, and cytochromes c1, c, a, and a3 (with prosthetic groups containing Fe or Cu)]
- then to O2 (forming water)
Follow the electrons in this diagram (ETC handout). The electrons from an NADH2 are transferred to FMN (a simpler form of FAD) and then CoQ, reducing it to CoQH2. Subsequently, the electrons are passed from CoQH2: that is, CoQH2 gets oxidized by passing the electrons to cytochrome b. The cytochromes contain heme as a prosthetic group, in which an iron oxidation-reduction occurs: Fe+++, +1 electron ---> Fe++. So now cytochrome B has the electrons. Cytochrome B gets oxidized by an iron-sulfur protein and that in turn gets oxidized by a heme in cytochrome C1. Its iron returns to ferric (Fe+++), while cytochrome C's heme group get reduced from the Fe+++ to the Fe++ state. Similar transfers occur between cytochromes C1, C, A, and A3. Finally, cytochrome A3 passes the electrons to molecular oxygen, which also picks up 2 protons to go with two electrons to form hydrogen atoms, so that the product of this reduction is H2O, which is the final resting place for these travel-weary electrons.
Note that the electrons become separated from the hydrogen ions early in this process (after FMN); after that we follow the electrons and let the protons join the general proton pool, for the moment.
An energy level handout shows the free energy changes associated with some of these electron transfers. Each transfer is energetically favorable, with some of the changes releasing much more free energy than others. It can be seen here that the 53 kcal per mole for the reaction between NADH2 and oxygen has been broken up into smaller packets of free energy changes. The free energy changes marked with an asterisk are those that have been found capable of generating a molecule of ATP from ADP. We will get to the mechanism of that generation a little later. You can also see that whereas NADH2 can generate 3 ATPs, FADH2 can only produce 2.
Thus CoQ and the cytochromes C, and NAD and FAD, are constantly shuttling electrons, picking them up originally from glucose-derived molecules and then delivering them elsewhere and then returning to pick up another load.
In the end, O2 receives the electrons. All the reduced forms of the oxidative cofactors (NADH2, FADH2) return to the oxidized state (NAD, FAD), having gotten rid of these electrons..
So: ..... we have no NAD or FAD loose-end debt any longer.
All debts have been paid. All the glucose carbon atoms have been converted to CO2, and the electrons from glucose have now all been delivered to O2 to form water.
To review electron transport try problem 5-9 and then 5-10.
So where does the ATP come in, after all this bother? If ATP is the energy currency of the cell, show me the money.
(mitochondria)
To understand how ATP is generated in this
process of electron transport, we must discuss the special structures in the
cell where all this electron transport and ATP generation takes place: the
MITOCHONDRIA.
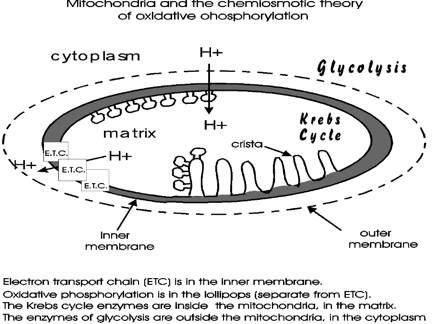
Some important features of mitochondrial structure can be seen in the simple diagram of the mitochondrion handout.
There is an OUTER membrane that is permeable to most small molecules (to MW 5000) and which need not concern us much here.There is an INNER membrane, with cristae, or extensive invaginations to increase membrane surface area, and which does provide a barrier to transport [Purves photo]
The reactions of the Krebs Cycle as well as the entrance reaction to the cycle take place in the inside of the mitochondrion, which is called the MATRIX. Glycolysis takes place in the cytoplasm: so the pyruvate produced must get in to the mitochondrial matrix. The ETC proteins complexes are held within the INNER MEMBRANE (within the cristae).The mitochondrion handout shows that these electron transport proteins of the ETC are organized into 3 groups; these protein complexes, called respiratory complexes are geographically fixed next to each other within a membrane in the cell, as we shall soon see. That is, these are membrane-bound complexes of proteins. [Purves picture].
But where is the energy? the ATP?
(chemiosmotic theory)
The answer lies in the
Chemiosmotic Theory first proposed by Peter
Mitchell in 1961.
(proton pump)
The energy released at each of the electron
transfers is stored in an electro-chemical gradient, established across the
mitochondrial inner membrane. Concomitant
with electron flow, H+'s are being pumped out of the matrix
into the inter-membrane space. These hydrogen ions are not clearly from NADH2
per se, as H+'s get pumped out even in later steps in
the ETC where no protons are directly involved (just electrons on Fe++ atoms)
[Purves picture]. So this pumping out
of H+ ions must be coupled the binding and release of electrons by the proteins
involved (see one proposed mechanism on the
handout).
The immediate effect of this H+ pumping is a higher concentration of H+ ions outside the mitochondrion's inner membrane and a lower concentration of H+ ions inside the mitochondrion. Now we allow the pumped-out H+'s to flow back. By mass action, the protons will flow from a region of high concentration to a region of lower concentration. A steady-state increment (between the outside and the inside of the inner membrane is thus maintained, resulting in the matrix (inside) being about 1 pH unit higher than the outside). A constant kicking-out, and flowing-back.
The H+'s may not get that far, but the outer membrane is no barrier to H+ ions, so how far they get is not an important factor.
To review the movement of protons and electrons see problem 5-13 part A ( part C-1 in 16th ed.)
(oxidative
phosphorylation and substrate level phosphorylation)
This process of forming ATP by an proton
motive force is called OXIDATIVE PHOSPHORYLATION
(OXPHOS).
{Q&A}. And so there are two methods of
producing ATP from glucose metabolism: Oxidative phosphorylation and the
"regular", direct phosphorylation of ADP from phosphorylated intermediates that
was seen during glycolysis, or in the GTP-forming step in the Krebs Cycle. This
direct phosphorylation is called SUBSTRATE-LEVEL
PHOSPHORYLATION (SLP), to distinguish it from OXPHOS.
Some evidence for the validity of the chemiosmotic theory of oxidative phosphorylation is:
1) Adding H+ ions (adding acid, in moderation) to closed vesicles (or membrane-bound spheres) that have been formed from membranes containing the F1-Fo protein complexes generates ATP from ADP + Pi in the test tube.
2) Isolated ETC complexes I, III, or IV inserted into artificial membranes are able to pump H+ ions in the predicted directions when provided with the appropriate substrates (reduced electron carriers like NADH2).
3) DINITROPHENOL, a small partially hydrophobic molecule, can return H+'s to the inside of the mitochondria via a short circuit; it ferries them across the inner membrane; the H+'s thus avoid the Fo channel. This compound uncouples H+ transport from ATP generation, so you get electron transport, but no OX-PHOS (since there is no longer a build-up of a proton gradient across the membrane).What about E. coli, they have no mitochondria (in fact a mitochondria is about the size of an E. coli cell). Bacteria simply use their own plasma (cell) membrane, and kick the H+'s out into the medium (or in the immediate vicinity of the periplasmic space between the cell membrane and the cell wall).To review all of oxidative phosphorylation try problems 5-11 and 5-12.
OK, so how much ATP do we get after all this?
1 ATP per pair of electrons transferred through EACH of the 3 enzyme complexes (I, III, and IV). The number of protons transferred per pair of electrons is not really known precisely (it's been estimated at 10-12, so 3-4 protons flowing back can produce an ATP).
So 3 ATPs per pair of electrons passing through the full ETC.
So 3 ATPs per 1/2 O2
So 3 ATPs per NADH2
But only 2 ATPs per FADH2, which skips complex I, and delivers its electrons to CoQ via complex II, with little free energy released at that first step.
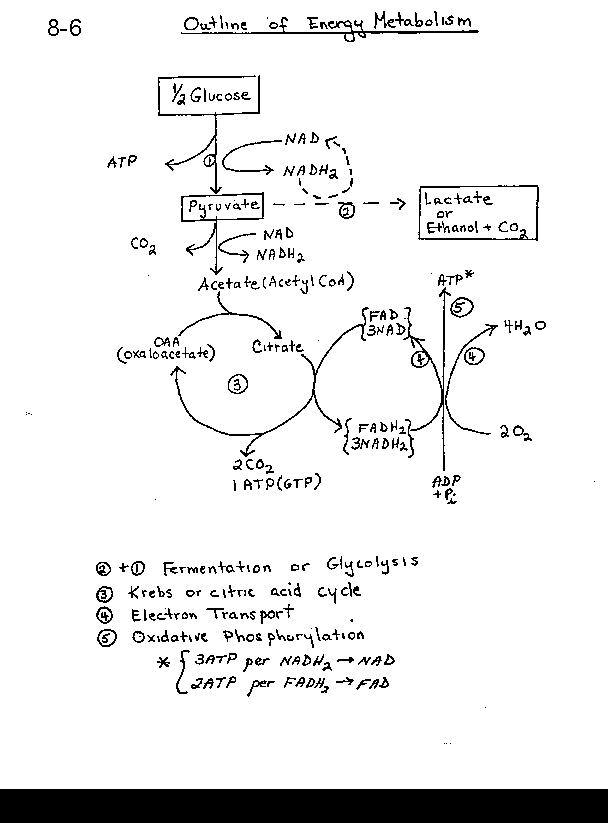
Overall ATP tally of RESPIRATION (as this overall oxidative metabolism of glucose is called) (see also OUTLINE of energy metabolism handout):
ATP from substrate-level phosphorylation, SLP (per glucose):Glycolysis: -2 that have to be invested, then +4 for a net of +2
Krebs Cycle: 2 (as GTP)
Total SLP = 4
ATP from OxPhos (per glucose)
(first calculating per Glyceradhyde-3-P, where the first oxidation takes place:
1 NADH2 from glycolysis and 1 from entry into the KC and 3 from the KC proper)
So 5 NADH2 @ 3 ATP/NADH2 = 15
FADH2: 1 from the KC @ 2 ATP/FADH2 = 2
Total OXPHOS per molecule of glyceraldehyde-3-phosphate = 17
Per glucose molecule, multiply by 2:
17 X 2 = 34
Grand total = 4 + 34 = 38 ATPs per glucose (in E.coli and other prokaryotes)
Now, if we are considering eukaryotic cells, we need to subtract 2 ATPs from this total, as 2 ATPs are used to get electrons from the 2 cytoplasmic NADH2's from glycolysis into the mitochondria (by an indirect mechanism), so the net for eukaryotes is 36.
Efficiency: 36 ATPs X 7 kcal/mole ATP hydrolysis = 252 kcal/mole harnessed as ATP.
252/686 available from glucose combustion = 37% efficiency.
Once again, better than most engines (20-25%).
And compare: 36 ATPs per glucose from respiration to 2 per glucose for fermentation . {Q&A}
So with or without air, ATP is no problem............
To review overall energy metabolism and compare fermentation and respiration, try problems 5-1, 5-3, 5-5.
To review energy (ATP) yields +/- oxygen, try problems 5-2 and 5-4.
© Copyright
2006 Lawrence Chasin and Deborah Mowshowitz Department of Biological Sciences Columbia University New York, NY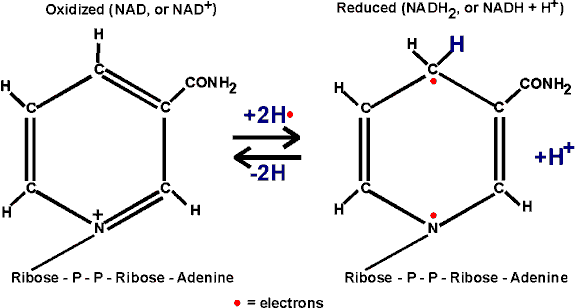{kind=link}
![[Purves picture]](../purves6/figure07-03.jpg){kind=link}
{kind=link}
![[Purves picture]](../purves6/figure07-07a.jpg){kind=link}
![[another Purves picture]](../purves6/figure07-07b.jpg){kind=link}
![[yet another Purves picture]](../purves6/figure07-07c.jpg){kind=link}
![[Purves picture]](../purves6/figure07-06.jpg){kind=link}
![[Purves picture]](../purves6/figure07-05b.jpg){kind=link}
![[another Purves picture]](../purves6/figure07-15.jpg){kind=link}
{kind=link}
![[Purves picture]](../purves6/figure07-09a.jpg){kind=link}
![[another Purves picture]](../purves6/figure07-09b.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![[Purves picture].](../purves6/figure07-11.jpg){kind=link}
![[Purves picture]](../purves6/figure07-13a.jpg){kind=link}
{kind=link}