{kind=link}
C2005/F2401 '06 Lecture #12 -- Wrap Up of DNA Synthesis; PCR; What is RNA & What is it Good For? Next time: How is RNA Made?
Handouts: 11-3 -- DNA Replication - Details at Fork & 12A -- PCR (Not on web.) 12B = code table and ribosome structure. (See texts.)
© Copyright 2006 Deborah Mowshowitz and Lawrence Chasin,
Department of Biological Sciences, Columbia University New York, NY.
Last updated
10/17/2006 01:36 PM
(A few minor typos fixed after the am lecture.)
References are to 6th ed. of Becker and 7th ed.
of Purves. (Refs in parentheses are to previous editions, 5th for Becker &
6th and 5th
for Purves.)
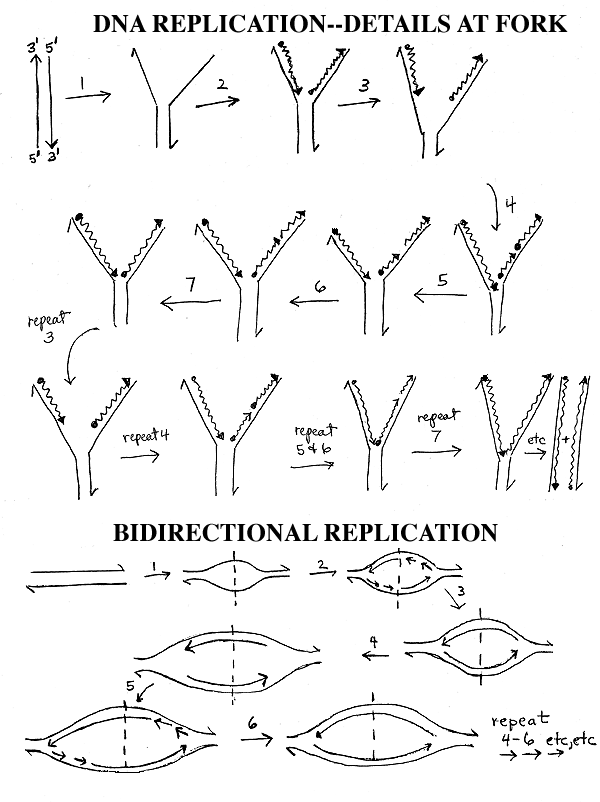
I. Discontinuous synthesis, cont. Topology.
In the pictures on the handouts and in many pictures in the texts, it looks like the DNA stays put and the enzymes move down the DNA. It is probably the other way around -- the enzymes stay put, and the DNA slides through them. See Purves fig. 11.11 & 11.12 (11.12 & 11.13 in 16th; not in 5th edition.)
FYI only: The polymerases making the leading and lagging strands are probably moving effectively in the same direction, relative to the fork, because the template for the lagging strand is looped around. See Becker fig. 19-14. (The leading and lagging strands are both being made in the 5' to 3' direction, anti-parallel to their respective templates.)
II. Bi-directional Replication. (Bottom
of handout 11-3).
A. How many replication forks per DNA? The more forks, the faster replication is. Most small genomes (such as bacterial and viral DNA's) are circular, and replicate bi-directionally -- 2 forks emanate from a single origin as shown on the bottom of handout 11-3 or Purves 11.11a & 11.12a (11.12a &11.13a in 6th ed., or 11.13 a in 5th ed) or Becker 19-5 (17-5). Longer DNA molecules are usually linear and often have multiple bidrectional origins of replication as shown in Purves 11.12 b (11.13b) or Becker fig. 19-6 (17-6) -- this will be discussed later when we get to eukaryotes.
B. How does bi-directional Replication go? In the top picture on the handout you have one fork or zipper moving down the DNA. In the bottom picture, you have 2 zippers or forks. Both start from the same point (the dotted line = origin of DNA replication = ori) but one fork goes to the left and one fork goes to the right. The events at each fork are the same as those shown in the top of the handout, but the forks go left and right instead of down. At each fork you have unwinding, continuous synthesis on one strand and discontinuous synthesis & ligation on the other strand, just as before. If the DNA is circular, the right fork is really going clockwise and the left fork counterclockwise, and the 2 forks proceed until they meet in the middle of the molecule, approximately 180 degrees from where they started. (See Becker fig. 19-5 [17-5].)
C. An Important Definition: Bidirectional replication means that there are 2 forks that move in opposite directions. It does NOT refer to the fact that the 2 DNA strands (leading and lagging strands) are made in opposite directions. That is called discontinuous synthesis, and it always happens at every fork whether there is one fork (unidirectional replication as in the top panel of handout 11-3) or two (bidirectional replication as on the bottom of the handout.)
To be sure you understand what is happening in the bottom picture, it is a good idea to write in all the 5' and 3' ends on the DNA's shown and also to number the Okazaki fragments at each fork to show the order in which they are made.
To review bi-directional replication, see problem 6-13, part A. (And be sure you can properly label the picture on the bottom of handout 11-3.)
III. Primers & Primase. (Top of handout 11-3.)
A. The Problem
If you put DNA polymerase, ligase, pyrophosphatase,
dATP, dGTP, dTTP & dCTP in a test tube (+ all unwinding enzymes) will you get DNA? No, because DNA polymerase can't start a new chain
-- it can only add on to the 3' end of a pre-existing chain. So how do new
DNA strands get started? Using primer and primase.
B. The Solution in vivo
1. How Primase makes Primer -- see Purves 11-14 (11-15 in
6th ed) or
Becker 19-11 (17-11).
a. Primase: Primase is a type of RNA
polymerase that uses ribonucleotide triphosphates to make a short RNA stretch (probably less
than 20 bases long {Q&A}) complementary to the 3' end of the template (= 5'
end of new strand). RNA polymerases (unlike DNA polymerases) can start new
chains from scratch.
RNA chains are made 5' to 3' in much
the same way as DNA, using ribo-nucleoside triphosphates instead of deoxy-nucleoside
triphosphates; details next time.
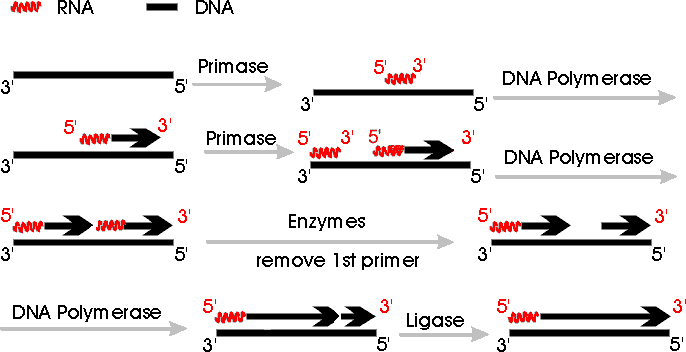
b. Primer: Short RNA stretch made by primase is called primer. On handout (11-3) of events at fork, RNA primer is represented by a dot. (In diagram below, primer is a red squiggly line.) Primase catalyzes synthesis of primer, and then DNA polymerase adds on the 3' end of the RNA primer.
2. How
Primer is Removed & Replaced
The primer (the short RNA section) must be
removed and replaced by DNA. The process is shown in steps 5 & 6 of handout
11-3 and in the diagram below. They are described separately to make the process
clearer.) Once primer is removed (step 5), there is a gap
between the 3' end of Okazaki fragment #2 and the 5' end of fragment #1. DNA polymerase
adds on to the 3' end of primer #2 to fill the gap (step 6), and then ligase joins the loose
ends (step 7).
Removal of RNA primer (step 5) and filling of the gap with DNA (step 6) may
occur simultaneously, using two different catalytic parts of a single enzyme.
3. Summary Pictures of Use & Replacement of Primer
See Becker Fig 19-13 (17-13) or Purves 11.17 (11.18 in 6th ed) or Picture Below. Note:
Some of the pictures in the older editions of the texts don't have all the details right.
Some of the figures imply that DNA can replace RNA primer
without the need for a free 3' end for DNA polymerase to add on to. Other
figures show ligase joining the Okazaki fragments at the wrong place. (See
picture below or solution to problem 6-14, part B-3, for correct position of
ligation. Note that the replication fork in problem 6-14 goes in the opposite
direction from the fork in the picture below.)
In the picture below, which summarizes the process of primer synthesis and replacement, all arrows go 5' to 3'. Only one side of the replicating fork is shown -- the side carrying out synthesis of the lagging strand. The side carrying out continuous synthesis is omitted. Note that replication fork below goes right to left -- DNA is unzipping from right to left.
Function & Replacement of Primer; see also handout 11-3.
C.
Telomeres and Telomerase -- a
Biological consequence of the need for primers
1. The "loose end" Problem
There is no easy way to replace the primer on the left end of the new strand (in picture above); also see Becker fig. 19-15 (17-14). The RNA can be removed, but no DNA can be made to fill in the gap.
2. Solutions
a. Small DNA's
(& most prokaryotic chromosomes) are generally circular, which circumvents
this problem.
b.
Linear chromosomes (the norm in eukaryotes) tend to get shorter with each
replication -- in next go round, chain that has just been made will be
template, and it is shorter than the original by the length of the primer. The
consequences will be discussed next term when we focus on eukaryotes.
FYI: Eukaryotic chromosomes get shorter with each replication, but it doesn't usually matter because the DNA molecules in the chromosomes have special repeated structures called telomeres on the ends. The repeats are gradually lost unless replaced by an enzyme called telomerase. See Purves 11.18 (14.3 in 6th or 14.5 in 5th ed.) or Becker 19-16 (17-15). (Note: in Purves' picture in older editions, the wrong strand is "too short." The 3' end should be longer than the 5' end, not the reverse.) The lack of telomerase may be what limits normal somatic cells to a finite life span of 50-60 divisions; the presence of telomerase may be what allows indefinite multiplication of cancer cells. Germ cells, that produce eggs and sperm, make telomerase, so a new generation always starts out with full length telomeres. For an animation of how telomerase works, see http://faculty.plattsburgh.edu/donald.slish/Telomerase.html. (Note that this is for eukaryotes; in this case there are two different DNA polymerase for the leading and lagging strands. Both grow chains in the 5' to 3' direction.)
To review primers, see problem 6-12,
A-D and problem 6-14.
If you have the 2004 ed. of the problem book, there is a typo in problem 6-12, part D .
The 2005 edition is corrected. For all
corrections in notes, current problem book, etc., see the
corrections page.) For corrections to the 2004 problem book, see the
17th edition corrections page.
IV. PCR (Polymerase Chain Reaction) -- Handout 12A. A Practical Application of the need for Primers.
The inventor, Kary Mullis, received the Nobel prize in 1993. For his acceptance speech, biography, etc. see the Nobel Prize official site. Also see class handout.
A. Idea of prefab primer, hybridization.
DNA synthesis will not start without a primer. In a living cell, primase (a type of RNA polymerase) makes the necessary RNA primer. Then DNA polymerase can take over, adding on to the 3' end of the primer. In a test tube, you can omit primase and use an oligonucleotide (short polynucleotide, usually DNA) as primer (= prefab DNA primer) to force replication to begin wherever you want. The primer you add will hybridize to its complementary sequence, wherever that happens to be (not necessarily at the end of the DNA) and DNA polymerase will add on to the 3' end of the primer, thereby starting elongation of a chain from wherever the primer is.
B. Steps of PCR -- see PCR handout (12A), Purves 11.20 (11.21 in 6th or 16.12 in 5th), and/or Becker Box 19 A (17A). For an animation, go to http://www.dnalc.org/ddnalc/resources/pcr.html
1. First Cycle: You denature your template and add primers (one to each strand). Then you cool the mix down, and each oligonucleotide primer hybridizes to its complement (step 1 on handout). The two long strands of template do not renature.) Then DNA polymerase adds on to 3' end of primer until it reaches the end of the template strand (step 2). This completes the first cycle. The new strands you just made (dashed on handout) include the target sequence, plus some extra DNA on their 3' ends. (This "extra" corresponds to the sequence between the target area and the 5' end of the template strand.)
2. Second Cycle: You heat the DNA to denature it (step 3), and add more of the same primers as before (step 4). Then you allow DNA polymerase to add on to the 3' ends of the primers (step 5). This completes the second cycle. On the handout, only the fate of the new strands made in cycle two is shown after steps 4 & 5. The old strands simultaneously go through another cycle just like the one above (steps 1 & 2), but this is not shown on the handout. The new strands you made in cycle 2 (at step 5) include only the target sequence.
3. Third Cycle: You heat the DNA again to denature it (step 6), add primers (step 7) and allow DNA polymerase to add to the primers (step 8). This completes the 3rd cycle. On the handout, only the fate of the new strands made in cycle two is shown after steps 7 & 8. (The fate of the complementary strands, left over from the previous cycle, is to repeat steps 4 & 5.) At the end of this cycle, you finally have double-stranded DNA molecules the length of the target sequence.
4. Additional Cycles: After each cycle you heat the reaction mixture to denature the DNA, and then you cool the mixture down to start the next cycle. In each cycle, primer sticks to the appropriate spot (its complement) and polymerase starts at the 3' end of the primer and goes to the end of the template. Note that primers are complementary to sequences in the middle of the original chain, but that after two cycles the parts beyond the primers are no longer copied. {Q&A}.
5. How reaction is actually carried out. All components (template and excess of heat resistant polymerase, primers & dXTP's ) are present from the very beginning. The mixture is heated and cooled repeatedly to end and start subsequent cycles. You don't have to add primers, polymerase, etc. to start each cycle. Note: New molecules of primer are used in each round. However, the primer molecules used in each round have the same sequences as the ones used in all the previous round. The primers are not reused -- new primers (with the same sequences as before) are needed for each cycle. You need only two types (sequences) of primer, but you need many molecules of each.
6. Identification of Product. The products of the PCR reaction are usually identified by their lengths, which are determined by gel electrophoresis. Gels are used that separate DNA molecules on the basis of their molecular weights (which depends on chain length). Hybridization to labeled probes is often used to detect the positions of the bands of DNA on the gel. (More on this later.) An animation of DNA gel electrophoresis is at http://www.dnalc.org/ddnalc/resources/electrophoresis.html
C. Special Features of PCR (as vs. regular DNA synthesis)
1. Special Polymerase. The DNA polymerase used in this procedure is a special heat-resistant one (called Taq polymerase) that is not denatured when the temperature is raised to separate the two strands of the DNA. This special polymerase was isolated from bacteria that live in a hot spring.
2. No discontinuous synthesis. Note that the entire template molecule is denatured completely before each cycle, so each strand can be copied continuously. There is no replication fork and thus no discontinuous synthesis here.
3. No RNA primer. Primase is absent, so no RNA primers are made. Oligonucleotides of DNA (not RNA) are added instead to act as primers.
To review the
PCR technique, see problem 6-13, C-1 and 6-15.
For an animation of PCR and links to animations of other DNA techniques,
click here .
(This is the site with the animations of PCR and
electrophoresis described above.)
D. Uses/Advantages of PCR
1. Amplification: Uses small number of starting molecules & produces large number of copies of target sequence.
The beauty of this scheme (PCR) is that the desired (target) sequence is copied exponentially and the other parts of the original DNA are copied linearly. So after a few cycles you have lots of copies of the target sequence (and not much of anything else). To convince yourself of this, see the answer to problem 6-13, part C-2. To use this technique and make many copies of the target sequence all you need (in theory) is ONE starting DNA molecule (and appropriate primers). Given current technology, you need 10-50 starting DNA molecules (see articles handed out in class). You can use the multiple copies for many different purposes such as characterization and/or identification as explained below.
2. Detection -- Can be Used to see if a particular target DNA is present or not.
You can add primers to a sample that you suspect contains some particular target DNA, such as HIV DNA, or DNA from genetically modified corn, or DNA from a species of protected whales. The primers are complementary to a sequence found only in the target DNA -- the one you are testing for. (In the cases mentioned, the primers would be complementary to a sequence in HIV DNA, or a sequence added to ordinary corn DNA by genetic engineering methods to make the special corn, or to a DNA sequence unique to the protected species of whales.) Then you see if polymerase can make DNA. If no target DNA is present, primers will have nothing to hybridize to, so polymerase will have nothing to add on to, and no copies of DNA will be made. So if you don't get multiple copies, it indicates there was nothing to copy -- your target DNA was not there. If you do get multiple copies, your target DNA was in the sample.
Notes: (1) The standard HIV screening test is not for HIV itself or for HIV DNA but for antibodies to proteins of HIV. (PCR may be used as a backup to confirm a positive result with the standard screening test, or to measure the actual levels of HIV.)
(2). Why would you test for genetically modified corn? StarLink corn is a type of genetically modified corn that was approved for animal feed, but not for human use. In spite of attempts to keep it separate, it has turned up in many human foods. It is probably harmless to humans, but no one wants to take any chances. Testing for the modified DNA is the only way to tell if StarLink corn is present in a mixture or not.
3. Forensics -- Can be Used for identification -- DNA fingerprinting
a. Basic idea: PCR can be used to copy specific sections of the DNA from different samples -- for example, from DNA left at the scene of a crime and from DNA from a suspect. The sections of amplified DNA can then be compared to see if they match or not (in length, sequence, etc.). The sections that are compared are highly variable ones that probably don't carry any information and are merely spacers in the DNA. If enough sections are checked, you can determine (to a very high degree of certainty) whether the two DNA samples came from the same person or not. DNA testing can be used to identify the guilty (inclusions) and to clear the innocent (exclusions). Alec Jeffreys, who first came up with the idea of using DNA testing for identifications, received a Lasker award in '05. For a pdf with details see the Lasker site.
b. Examples: See articles handed out in class and article from the San Francisco Chronicle of 10/19/99. (Note: you'll need to go to the SFChronicle web site itself if you want to see the pictures or get some of the older articles.)
c. Inclusions: If the samples match at enough highly variable spots, then there is a very high probability the samples came from the same person, because the degree of variation is so high that only a few different people in the world should have the same pattern.
d. Exclusions: If the two samples do not match, then it is clear that the two samples came from different individuals and the suspect could not have committed the crime (since the DNA at the scene came from someone else).
e. STR's: The variable sections that are tested are often ones that have different numbers of short tandem repeats (STR's). The primers hybridize to regions outside the section with the repeats. The number of repeats in each DNA can be figured out from the length of the sections amplified by PCR. The new FBI data base contains the information from checking 13 sections with variable numbers of STR's.
For a great site with examples of how DNA is used for identification and forensics click here.
4. Why you can't do this with proteins
There are very sensitive tests for presence of proteins (usually using the catalytic activities of enzymes and/or binding abilities of antibodies), but no way to amplify (make copies of) what you detect. you can't make more. PCR takes advantage of fact that DNA replicates for a living to make more copies.
Note: So-called DNA fingerprints are characteristic of the person/DNA from which they came. So-called protein fingerprints are characteristic of the protein from which they came. That's why both are called 'fingerprints.' However the two types of 'fingerprints' are made differently and used for different purposes.
V . Central Dogma -- How does DNA do job # 1?
A. Big Picture. So we have a big DNA that includes a particular gene = stretch of DNA coding for a single peptide; how will we make the corresponding peptide?
Note: gene usually means a stretch of DNA encoding 1 polypeptide, but there are complications as we'll see later.
Basic idea -- see also Becker fig. 21-1 [19-1] or Purves 12.2 & 12.3:
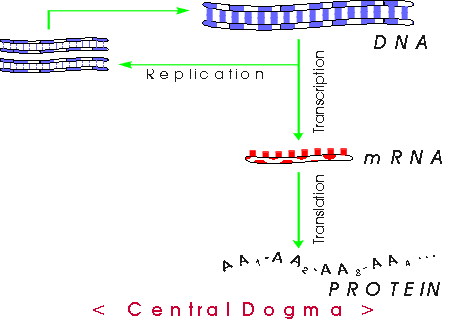
Transcription = RNA synthesis using DNA template; translation usually = protein synthesis using RNA template, but can mean entire DNA --> RNA --> protein.
B. What RNA is
1. Structure: See Purves fig. 3.25 (3.17 in 6th or 3.22 in 5th) and table 3.3 or Becker table 3-5 (3-4) & fig. 3-17 for comparison of DNA and RNA. RNA is single stranded (although sections may double back on themselves → double stranded regions), has U not T, ribose not deoxy and is generally shorter, but otherwise like DNA. RNA is less stable than DNA -- more easily damaged (because of reactive OH on ribose and because a single strand is more exposed) and less easily repaired (because no 2nd strand to use to correct mistakes on first strand). DNA is also more easily repaired because it has T not U, so damaged C's (which are oxidized to U) can be recognized and removed. In summary:
| DNA | RNA | Significance/Effect of Difference |
|
Double Stranded |
Single Stranded |
|
|
|
U not T |
For DNA: Ease of repair of damaged (oxidized) C up. (Damage that coverts C to U can be detected & repaired.) |
|
|
Ribose |
For RNA: Reactivity up, stability down |
|
Very long |
Relatively Short |
|
2. Synthesis. RNA grows just like DNA by adding XTP's and using an enzyme called RNA polymerase; details to follow.
VI. Why mRNA?
A. Basic idea: mRNA = Working, disposable copy vs DNA = archival, permanent master copy. DNA = big fat comprehensive reference book or complex web site. mRNA = Xerox of one (book) page or print out of one web page with information you need for a particular assignment. Book stays safe in library; web site remains unchanged. Xerox goes to your room, is actually used, gets covered with coffee stains, smudged, and thrown away.
B. How function of mRNA corresponds to structure
1. Convenience -- small size (1 or a few peptides' worth) is much more convenient than many genes' worth. Xerox of one page much more convenient to work with than big fat book.
2 . Flexibility in different rates of synthesis of different proteins. Cells can vary the amount of mRNA made/gene so they can vary the rates for synthesis of different proteins . If mRNA is template for protein synthesis, a cell can make many mRNA copies of DNA template for protein B if it needs to make lots of protein B. If DNA were direct template, cell would have only 1 copy of template no matter how much protein B it needs to make. (If several people are working on the same assignment, you need more Xeroxes.)
3. Flexibility in response to needs that change with time -- bacterial cells constantly make new mRNA's, and degrade old ones (to mononucleotides) so a cell can constantly adjust the rates of protein synthesis (and change which ones are made) to meet the current need. DNA has a long half life, as expected of a master copy ; bacterial mRNA has a short half life, as expected for a working copy. (When assignment is done, you crumple up the Xerox and throw it away. Then you make a new Xerox for the next assignment.) See Becker, Key Aspects of mRNA metabolism, p. 680 (654) at end of Ch. 21 (19).
Note: In eukaryotes the situation is more complex -- some eukaryotic mRNA's last a long time while some are short lived. It depends on the cell type and what protein the mRNA codes for.4. Preserve Master. Using mRNA to make protein saves wear and tear on master -- no coffee stains on the archival copy (DNA).
VII. Why other kinds of RNA for Translation? This will be covered in lecture 13.
A. How decode mRNA (general idea)
1. It's read in triplets . Reading starts at a fixed point and then mRNA is read one triplet or codon at a time in the 5' to 3' direction.
2. Code table (see texts or handout) lists codons = triplets found in the mRNA (NOT complements of codons) and corresponding amino acids. One codon specifies one amino acid. For example, CUA means leucine; UUU means phenylalanine, AUG means methionine.
B. What is tRNA & what is it for?
1. Need an adapter -- how does cell know AUG is met and CUA is leu? You have the text or handout with the code table, but cell doesn't.
a. Transfer RNA (tRNA) = adaptor. Cell uses tRNA to match the codon in the mRNA (say AUG) which specifies the amino acid (say met).
b. Loading Enzymes. Adaptor must carry the correct amino acid. Cell uses loading enzymes to put the correct amino acids on to their respective tRNA's. More details next time.
2. Properties of tRNA
a. Size: About 75 bases long (relatively small).
b. Many different ones . Actual number of dif. tRNA's is more than 20 (#of dif. amino acids) and less than 64 (# of dif. codons). More exact estimate of # of different tRNA's to follow in next lecture.
c. Two headed molecule: tRNA has 2 ends (in its 3D form) -- one complementary to codon (= anticodon) and one (on 3' end) to pick up the appropriate amino acid (= acceptor end) with the help of the appropriate enzyme.
d. General features of structure -- every tRNA molecule is folded back on itself to form some double stranded regions. Sequences of different tRNA's differ, but all are self complementary in certain regions. Every tRNA molecule has same basic "secondary structure" = cloverleaf; this is folded into an L shaped "tertiary" structure, which has anticodon at one end and acceptor for its amino acid at the other. (See Becker fig. 22-3 [20-3], or Purves 12.7 (12.6), for secondary and tertiary structures.)
C. What is rRNA & what is it for? Ribosomes and rRNA
1. Function. You need something to hold tRNA (two loaded ones at a time) onto mRNA while amino acids are being hooked up and you need to provide necessary enzymes for making peptide bond etc. (How many weak bonds hold a tRNA and mRNA together?)
2. Structure. Holding of tRNA etc. is done by a structure that contains both RNA(s) and protein(s). Anything made of both is called an RNP = ribonucleoprotein or ribonucleoprotein particle. This particular RNP structure = ribosome; RNA inside it is called ribosomal RNA or rRNA. So need rRNA as well as mRNA and tRNA. For pictures of ribosome structure (details next time) see Purves 12.9 (12.8) and/or Becker figs. 22-1 & 22-2 [20-1 & 20-2] & table 22-1 [20-1]. )
D. Summary: How does RNA make protein?
1. "RNA makes protein" means two things:
a. Need mRNA (info goes DNA → RNA → protein) -- see Purves 12.2
b. Need several kinds of RNA to make each protein: one mRNA, tRNA (at least 30 dif. kinds) and rRNA ( 3-4 kinds). Of course you need protein too to make protein.
2. Hardware vs. Software. rRNA and tRNA are the hardware or tools or machines; mRNA is the software or working instructions or tapes/CDs/punchcards. Cells use same old hardware and constantly changing, up to the minute, supply of new software.
Next Time:
Where does the RNA come from? So you need lots of kinds of RNA. How do you make them? All RNA is transcribed from a DNA template. See Purves 12.4 or Becker fig. 21-9 & 21-11 [19-9 & 19-11]. The easiest way to go over RNA synthesis, given that we've discussed DNA synthesis at length, is to compare DNA and RNA synthesis. We'll go over this next time, and then consider how the RNA is used to make protein.
© Copyright 2006 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY