C2005/F2401 '06 -- Lecture 15
-- Last Edited: 10/29/06 12:20 PM
© Copyright 2006
Handouts: 15A -- Induction vs Repression; Repression vs Feedback Inhibition & 15 B -- Operons
I. Mutations. (This is a repeat of Topic VI of Lecture 14)
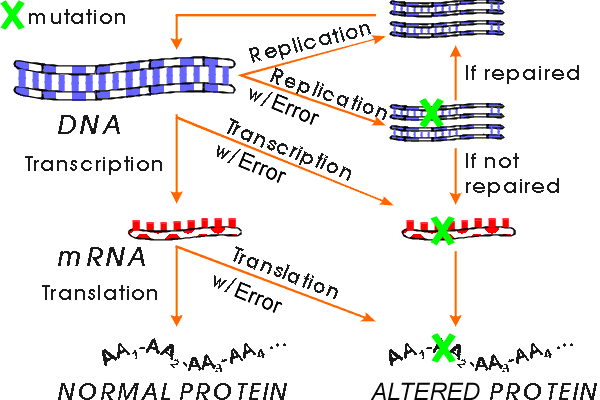
A. What are consequences of mistakes in macromolecular synthesis?
B. How do mistakes in DNA synthesis occur?
Bases can mispair (if in wrong tautomeric form or damaged -- See Purves 12.19). DNA polymerase can slip relative to the template and add extra bases or leave some out. Proofreading keeps mispairing mistakes low but not zero. Repair enzymes fix some mistakes. (see below for why mutations are necessary) After a mistake (putting in wrong base) occurs, other strand is still okay. But if DNA with mistake in one strand is replicated before mistake is fixed, one of daughter molecules will have two changed strands. (Other daughter molecule will be okay.)
C. Definition/terminology of mutations See Becker Box 22B p. 700 (20B p. 685) or Purves pp. 250-252 (234-235).
1. Mistakes vs Mutations. Mistakes in RNA or protein synthesis are called mistakes, but mistakes in DNA synthesis (that are not corrected) are called mutations. Anything that changes the DNA is called a mutation, and an organism with a mutation is called a mutant. The normal or starting (or standard) organism is often called "wild type." A change in the RNA or protein that does not affect the DNA is not called a mutation.
2. Substitutions vs deletions/insertions/frameshifts.
Substitution = change in base(s); deletion/insertion = removal or addition of base(s). An insertion/deletion of 1 or 2 bases is called a frameshift because mRNA with such a mutation is misread in the wrong "reading frame" (wrong groups of three nucleotides) all the way to the end of the gene (or until ribosome reaches a stop codon). Notice the drastic difference in effects between substitutions and frameshifts. A mutation that generates a stop codon is sometimes called a "nonsense" mutation; one that changes one amino acid to another is called a "mis-sense" mutation. See Becker Box 22B (20B) or Purves p. 252 (276).
3. Phenotype and genotype
The state of the DNA is known as the genotype; the observable properties of the organism are known as the phenotype. A mutation changes the genotype, but may or may not change the phenotype. See recitation problems #8.
D. Why are mutations important?
1. Source of evolutionary diversity -- source of all variation in phenotype for selection to act on; why there are different species (& why we're here at all). This is good overall, but not good for us when it's HIV or flu or any other infectious agent that's mutating.
2. Source of individual (& nonfunctional) diversity. Mutation leads to variations in noncoding DNA. This has little or no functional consequences, but the variations come in handy for tracing evolutionary lines of descent and making identifications. (This is the basis of all forensic ID's.) Variations that do not affect phenotype persist because there is no selection for or against any particular version. (Individuals that carry any particular mutation are not at any reproductive advantage or disadvantage.)
3. Cause inherited diseases like hemophilia, Tay Sachs, etc. (Can cause cancer in somatic cells.) To keep advantages of (1) and avoid disadvantages of (3), organisms keep the mutation level low but nonzero by extensive editing, repair, etc. of DNA
4. Mutations are a very useful tool for figuring out how things work.
- Studying effects of frameshifts allowed us to start to crack the genetic code -- see prob book, recitation problems and texts (for ex. Becker pp. 655-657 [640-643])
- Allows us to knock out one protein at a time and see what happens -- implies what function of protein was in first place. (As in figuring out pathways in prob. book.) Same affect can often be achieved with RNAi (or antisense).
Note: In this course, we often tell you how it works first and then give you mutations to test your understanding. Historically, it usually works in reverse -- mutations are studied first and details of how it works are figured out from analyzing the mutants. For example, the genetic code was partially "cracked" by looking at mutations and seeing how changes in the DNA correlated to changes in the corresponding protein. (It took biochemistry to finish the job.) See texts for details.
To review mutations, see recitation problems #8 and problem 7-22. (7-23, 7-24 & 7-26 also deal with mutations.)
II. Intro to Regulation in Prokaryotes (See handout 15A)
A. Why regulation of enzyme synthesis is reasonable and/or necessary -- consider some typical enzymes -- glycolytic enzymes, beta-galactosidase (needed to breakdown and metabolize lactose = dimer of glucose and galactose), and trptophan synthetase (needed to synthesize trp). (See Becker 23-1.) When are these enzymes needed?
1. Glycolytic enzymes -- always needed
2. Beta-galactosidase -- only needed if lactose present (and needs to be broken down); enzyme level should be low until lactose added to medium.
3. TS (trp synthetase) -- only needed if trp low or absent (then trp must be synthesized in order to make proteins) -- enzyme level should be high until trp added to medium.
B. The Phenomena -- Are enzymes (like those above) actually made only when they are needed? Graphs on handout 15A show what happens to level of appropriate enzyme if you add, or take away, the appropriate small molecule, namely lactose (lac) or tryptophan (trp).
1. Example of Induction -- Lactose (small molecule) = inducer = signal to turn on synthesis of appropriate enzyme ; synthesis of beta-galactosidase (enzyme) is called inducible; phenomenon is known as induction. (See also Purves 13.13)
2. Example of Repression -- tryptophan (small molecule) = co-repressor = signal to turn off synthesis of appropriate enzyme ; synthesis of trp synthetase (enzyme) is called repressible; phenomenon is known as repression.
3. Constitutive synthesis -- Synthesis of some proteins, such as enzymes of glycolysis, is called constitutive = synthesis of enzymes is "on" at all times.
C. Summary of Terminology -- see table in middle of Handout 15A
Regulation is covered in problem set 12. To review the material in parts A-C, see Problem 12-1, parts A & B.
D. Comparison of Repression to feedback. Why do you need both types of regulation? Factors to consider:
Overall, have coarse control (repression/induction) vs fine control (inhibition/activation). See chart and picture on bottom half of handout 15A. See also Purves 13.14. Note: Enzyme activation and induction can be compared in a similar way -- Activation increases enzyme activity while induction turns on enzyme synthesis
After you have figured out the mechanism of repression, and want to review the differences between repression and feedback inhibition, try problem 12-2, esp. part D, and 12R-4. (Be sure to figure out repression before you try these problems -- do 12-1 C & 12-2 A-B first.)
III. Mechanism of Prokaryote Regulation (See handout 15B) -- Operons
A. How is co-ordinate control achieved? Upper Left Panel on handout -- idea of cluster or operon. (See Purves 13.16 or Becker fig. 23-3.)
1. Genes regulated together are linked -- genes to be controlled co-ordinately (turned on and off together) are next to each other on the DNA.
2. Polycistronic mRNA. The linked genes are transcribed as a unit to give one single mRNA . One mRNA is made per operon (not one mRNA per gene), because all the genes in a cluster share a single promoter. An mRNA able to code for several peptides (mRNA that comes from several genes) is called polycistronic (cistron = another term for gene).
3. Transcriptional Control -- Regulation is at level of transcription. The level of translation is controlled by regulating the synthesis of mRNA. This is the usual method for regulation of protein synthesis in prokaryotes.
Since mRNA has a short half life in prokaryotes, regulating mRNA synthesis controls the steady state level of mRNA. Translation per se (and degradation of mRNA) are not regulated here. (In some prok. cases and many euk. cases, these are regulated too.)4. Definition of an operon = group of linked structural (enzyme coding) genes and regulatory sites that are transcribed as a single unit. (Note: gene for repressor protein is sometimes considered part of the operon and sometimes not. Usually clear from context; role of repressor will is discussed further below.)
5. Punctuation. Note that translation and transcription have different stop and start signals. Transcription starts at promoters; translation at start codons (AUG). What are the consequences?
a. mRNA has UTR's. It has leaders (untranslated region on 5' end before first AUG or 5' UTR) & trailers (untranslated 3' end or 3' UTR).
b. Numbers: Number of transcription starts (Promoters) for a message is one; number of translation starts may be many (one per peptide) in prokaryotes.
c. Translation of a polycistronic mRNA starts at multiple start codons. A ribosome assembles at the first AUG and starts translation. After each peptide is completed, the ribosome may continue down the mRNA to the next start codon and start a new peptide chain. Alternatively, the ribosome may detach (and disassociate into subunits) when it comes to a stop codon. In that case a new ribosome forms at the next start codon and starts translation of the next peptide.
B. How transcription of cluster is turned off -- Upper Right Panel of 15B -- Role of Repressor & Operator -- operon that is "off" (See Becker fig. 23-4, top panel or Purves fig. 13.17 top panel.)
1. Role of operator (O) = DNA site to act as part of on/off switch -- binds repressor protein when repressor is in appropriate or active form (rectangle on handout).
2. Role of repressor protein = other half of on/off switch (with O). Repressor binds to operator and prevents RNA polymerase from binding to DNA and transcribing the operon. (Purves fig. 13.15)
a. There is a different repressor protein for each operon. Repressor binds to specific sequence of DNA found in its respective operator.
b. Synthesis of repressor protein is constitutive -- always on. Gene for repressor gene has a promoter but no operator. (State of repressor protein varies, not the amount; see below.)
C. How induction and repression occur -- Role of Effectors
1. Repressor protein is allosteric (has two forms) -- one that sticks to the operator and blocks transcription (rectangle on handout) and one that doesn't (round on handout). See Becker Fig. 23-5 (21-5).
2. Repressor binds effector (inducer or co-repressor). Each repressor/regulator protein is unique in that it binds the proper co-repressor or inducer (see below) as well as the proper operator.
3. Effector determines which form the repressor is in. The amount of repressor protein present doesn't change (see above); the form repressor is in does change. The small molecule effector (inducer or co-repressor) shifts the balance between the two forms thus shifting the equilibrium between free and bound repressor and turning the operon "on" or "off."
4. How does repressor get on or off the DNA? The picture on the handout implies that the repressor is either "on" or "off" the operator. There is actually an equilibrium between free and bound "sticky" repressor -- "rectangular" molecules are spontaneously coming on and off. The effector shifts that equilibrium, by binding to free repressor, and changing the affinity of the repressor for the operator. Or you can think of the effector as changing the relative concentrations of free rectangles and circles. (The direction of the change depends on whether it's an inducible or repressible operon -- see below.)
D. An example of Induction-- (see middle panel of handout 15B or Becker fig. 23-4 (21-4) or Purves 13.17). For an animation try http://vcell.ndsu.nodak.edu/animations/lacOperon/index.htm. (There are multiple animations on the web; if anyone finds one they especially like, please tell Dr. M. This site has multiple animations of biological processes.) For an animation with a different slant, try http://trc.ucdavis.edu/biosci10v/bis10v/media/ch10/lac_negative.html This goes into some extra fine points, but is clear and interesting.
What are the characteristics of an inducible Operon?
Effector molecule (inducer) that binds to repressor protein prevents repressor from binding to operator -- decreases supply of free rectangles by converting them to circles.
Effector (Inducer) shifts following equilibrium to right:
"Rectangle form" of rep. protein ("sticky" form that binds to O) ↔ "Circle form" (form that doesn't bind to O)
Empty form of repressor protein (without effector) sticks to operator.
E. Constitutive Mutants & Plasmids
1. What happens if repressor protein is mutant and doesn't bind to DNA at all? Operon gets stuck in the "on" position and the operon (and synthesis of enzymes from structural genes) becomes constitutive. This is an example of "negative control" -- the regulator/repressor protein is needed to turn the operon off. Doesn't matter if operon is inducible or repressible -- same thing happens!
2. What happens if operator is deleted? Is it the same as above?
See
problem 12-3.3. How do you test out the properties of constitutive mutants? Many experiments and problems involve having a cell with two copies of an operon. How is this possible? A bacterium has only one DNA molecule (chromosome) with one copy of each gene or operon.
Answer: Bacteria can carry mini-chromosomes called plasmids that have 'extra' genes. The 'extra' genes can be additional copies of the genes already in the cell. Therefore a bacterium with a plasmid can have two copies of a gene or two copies of a whole operon -- the bacterium can have one copy on its normal chromosome and another copy on a plasmid. Such a cell is called a partial diploid (see below.) The two copies do not have to be exactly the same -- one can be normal and one mutant, or they can both be different mutants. For example, suppose a bacterium has two copies of the lactose operon. Suppose one copy is constitutive and the other is inducible, or suppose both are constitutive. What should happen when you put the two operons together? Will both be constitutive? Both inducible?
4. Use of Mutants. Study of the properties of constitutive mutants was how induction/repression was figured out by Jacob and Monod, who received the Nobel prize in 1965 for their work. Now you can try it the other way -- you can use your knowledge of operon function to predict the properties of mutants, both singly and in combination. See Chap. 12 of the problem book.
5. Terminology
a. Haploid = A cell (or organism) with one copy of each chromosome. Therefore one copy of each gene. Example: bacteria.
b. Diploid = A cell (or organism) with two copies of each chromosome (usually one copy from each parent). Therefore 2 copies of each gene. Example: mammals.
c. Partial Diploid = A cell (or organism) that is basically haploid, but has two copies of a few genes. The 'second copy' of the few genes is usually on a plasmid. Therefore the partial diploid has one copy of most genes, but two copies of the genes on the plasmid -- one copy on the plasmid and one on the chromosome. (See below and/or next time for how you get a bacterium with a plasmid.)
To learn how to tell the types of constitutive mutants apart, see problems 12-4 & 12-8 & Becker table 23-2 (21-2).
F. Induction vs. Repression
1. What are the characteristics of a Repressible Operon? (See Becker fig. 23-6 (21-6) or Purves 13.18)
Effector molecule (co-repressor) that binds to repressor protein promotes repressor binding to operator -- increases supply of free rectangles by converting circles to rectangles.
Effector (co-repressor) shifts following equilibrium to left:
"Rectangle form" of rep. protein ("sticky "form that binds to O) ↔ "Circle form" (form that doesn't bind to O)
Full form of repressor protein (= effector-protein complex) sticks to operator.
2. How does this compare to an Inducible operon? Compare & Contrast to D above.
It may help
to make a table for yourself comparing induction & repression. Some questions to
consider:
(1) Which form, empty or full, sticks to the DNA?
(2) When
protein is made, is it sticky?
(3) How (in which direction) does the effector shift the equilibrium
between sticky and non-sticky forms?
3. Reminder: The repressor protein of each operon is unique, and binds only to its respective operator (& effector). It is important to remember that not all rectangles (or circles) are the same. Each unique repressor protein is allosteric and has 2 forms -- "sticky" and "nonsticky." For comparison, on the handouts, all "sticky" forms are drawn as rectangles and all "nonsticky" forms are drawn as circles. However, each repressor protein is different. The only thing all 'rectangles' have in common is that they all stick to their respective operators.
To review how operons work, do problems 12-1 and 12-2 A-B. To compare repression and induction, do 12-2 C, and 12-7.
G. Strong & Weak Promoters -- all promoters are not the same.
1. All Promoters are similar in structure and function -- all P's have to able to bind RNA polymerase and serve as signals to start transcription.
2. P's can be strong or weak
a. Weak Promoter --> little RNA polymerase binding --> low levels of transcription --> low levels of corresponding protein.
b. Strong Promoter --> lots of RNA pol binding --> high levels of transcription --> high levels of corresponding protein.
c. Why does strength of promoter matter? The strength of the promoter determines how much mRNA can be made. Actual amount of mRNA made at any time depends on both strength of promoter and extent of repression or induction.
3. Example of strong vs. weak Promoters: P of lac operon vs P of lac repressor gene
a. Promoter of lac operon is strong. P of lac operon = P for the structural genes; controls production of polycistronic mRNA --> enzymes for metabolism of lactose. Since this P is strong, you make a lot of mRNA and a lot of the corresponding enzymes.
b. Promoter of lac repressor gene is weak. P of lac repressor = P for the R gene; controls production of mRNA for lac repressor --> lac repressor protein. Since this P is weak, you make only a little of the mRNA, and relatively little of the repressor protein.
c. Why does this make sense? You need a lot of the metabolic enzymes (if you are growing on lactose as a carbon and energy source) but relatively few molecules (100 or so) of repressor protein.4. Note difference between Roles of O (operator) and P (promoter). P determines what the maximum level of transcription is; O (plus Repressor) determines what percent of maximum is actually reached (per culture, not per cell).
a. O (by binding to repressor) determines to what extent operon is "on" -- is operon running at full throttle or is it only partially turned on (or completely off)? We usually described operons as "off" or "on." Operons can be partially turned on if there is an intermediate level of co-repressor or inducer, so that part of the repressor protein is in the "rectangle" form and part is in the "circle" form.
Note: one molecule of repressor (in the "rectangle form") is not enough to shut down one operon. There has to be more than one molecule of repressor protein per operon to be sure each operator is always occupied with a repressor protein molecule.
b. P determines the maximum level of transcription = level when operon is fully "on" and running at full throttle.
H. Regulation in general. This will not be discussed in detail in class but is included here as a summary and to help you get the big picture. It will be discussed in detail next term when we get to regulation of eukaryotic protein synthesis.
1. All models of regulation are based on knowledge of operons. Why? Because operons were the first systems of regulation of protein synthesis to be understood.
2. Features of operons to consider
a. Transcriptional Control. The level of protein synthesis is controlled by controlling the level of transcription of the gene coding for the protein. The production of mRNA is the only step that is regulated. There is no direct control of translation -- no control of use or degredation of mRNA.
b. The 2-Part Switch. There's a switch (controlling transcription) with two parts -- A DNA sequence or site (the operator) and an allosteric protein (repressor) to bind to the site.
c. Negative Control. Regulatory system is "negative" -- meaning a protein (repressor) must function properly to turn transcription of the system off. If the repressor protein is missing or does not work, transcription is stuck in the "on" position.
d. Co-ordinate Control. Genes coding for proteins of related function are controlled together -- the level of synthesis of the corresponding proteins is co-ordinated. In operons, this is done by having all the (structural) genes involved clustered -- the genes are next to each other on the DNA and are conrolled by a single promoter --> single poly-cistronic mRNA.
3. Are these features universal? Does Regulation of protein synthesis always work the same way? Is what is true of E. coli true of the elephant? (Monod liked to think so.)
a. Transcriptional control is common. It is the primary way, but not the only way, to regulate protein synthesis.
b. Two part switches, consisting of a protein and DNA site are very, very common. The situation is often more complex than the one described above, especially in eukaryotes. There are often multiple sites and/or multiple regulatory proteins (which can interact with each other as well as with DNA) that can affect transcription of a particular gene. Details will be discussed next term.
c. Negative control is not universal. It is very common in prokaryotes; positive control (where a protein is needed to turn ON a gene) is more common in multicellular eukaryotes.
d. Co-ordinate control is common, but the mechanism is different in different organisms. Genes of related function are generally clustered in prokaryotes, and share a common "switch (P, O etc.)." Genes that code for multiple enzymes of the same pathway are generally NOT clustered in eukaryotes. Since each gene in the set is located in a different place, each gene has its own "switch." (But all the switches are tripped co-ordinately.) So there is generally no poly-cistronic mRNA in multicellular eukaryotes.
IV. How is bacterial DNA passed on?
A. Introduction to cell division -- How does 1 cell make 2?
1. How do you double cell contents? Consider the central dogma -- we've covered it all -- how to double DNA, RNA and protein, and how to regulate protein synthesis. Once you double the protein (enzymes), that allows doubling of everything else, like carbos, lipids, etc. So suppose you double everything in the cell. How do you get 2 cells from 1?
2. Why distribution of DNA is the critical issue -- Making two cells from one comes down to "once the program is doubled, how are the two copies distributed to daughter cells?" Stuff that is not part of the program (genetic material) need not be divided exactly, but because of the chicken and egg problem, there must be some in each daughter cell. (Need some ribosomes, RNA polymerase etc. in each cell. But as long as you have some, and the genetic material, you can always make more ribosomes, enzymes etc.)
B. How do prokaryotes do it? binary fission -- regular segregation of circular chromosome attached to membrane
1. What does the DNA (genetic information) of a bacterium look like? Each bacterium has one, circular, double stranded DNA molecule = chromosome; the chromosome is attached to the cell membrane.
2. How the Chromosomal DNA is distributed.
a. To start, you have one cell with one double stranded DNA circle attached to membrane.
b. DNA replicates by birectional DNA replication (two forks start from a single origin) --> two double stranded circles, both attached to membrane. (See Becker fig. 19-5 (17-5))
c. Circles grow apart as membrane is laid down between the attachment points of DNA to membrane --> two circles pushed to opposite ends of cell. (There is also an active process, other than growth of membrane, that pushes the two origins of DNA replication apart. This has only been recently discovered.)
d. To end, you need only to lay down a membrane (and wall) between the two halves of cell, each containing one circle (= complete double stranded chromosome). This → 2 complete cells.
e. Note this is not mitosis OR meiosis; it is a different process (binary fission). Mitosis and meiosis occur only in eukaryotes; they will be discussed later.
f. How will the genetic material in the two daughter cells compare? If there are no mutations it will be the same, and all descendants will be identical. All the descendants produced in this way (by asexual reproduction of a single founder) are called a clone. (Doesn't matter if "founder" is a cell, molecule, or organism.) Is there any way (besides mutation) to get new combinations of genes? To mix genes from separate clones? That requires bacterial sex.
Next time: How do bacteria & viruses have sex? How are the results of bacterial and viral crosses (i.e. sex) analyzed by complementation and recombination? (There will be a handout on all the details.)
© Copyright 2006 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY.