{kind=link}
{kind=link}
C2005/F2401 '06 Lecture 20 -- Meiosis, Life Cycles, & Introduction to Genetics
© Copyright
2006 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY.Last update 11/19/2006 03:33 PM. Handouts: 20A & B ( 20B = Super cycle and Nondisjunction. ) Also need 19A & B.
0. Wrap up of Karyotypes -- Chromosome Banding & Rearrangements
I. Overview of Meiosis
A. What is meiosis for?
1. Need for meiosis/reduction division -- to keep karyotype & ploidy constant from generation to generation.
Most of the cells of most higher organisms are diploid. Humans, for example, have 46 chromosomes, or 23 pairs, in virtually all of their cells. If eggs and sperm also have 46 chromosomes, the next generation, formed from the fusion of an egg and a sperm, would have 92 chromosomes. But clearly the chromosome # does not double each generation. So the eggs and sperm, unlike all other cells, must have only 23 chromosomes and be haploid. So there must be a way to make haploid cells from diploid cells. There is, and the process is called meiosis. During meiosis, one chromosome from each pair is picked at random so that the resulting haploid has 23 chromosomes instead of 23 pairs. Then 2 such haploids fuse, during fertilization, to give you back a diploid with 23 pairs.
2. Why bother with all this? Why sex?
After all, you could start the next generation with one complete diploid cell from either parent and save yourself a lot of trouble! Some organisms do reproduce this way, at least some of the time, but most organisms engage in sexual reproduction. They probably do so because each cycle of meiosis, followed by fusion, allows for a new combination of chromosomes. (Crossing over, which occurs at meiosis, also allows for new combinations of genes within chromosomes as well.) So it looks like sexual reproduction is useful because it allows reshuffling of the genetic material (same argument as for bacteria). Reshuffling is needed to give
new variety (for selection to act on) and/or for repair (& replacement) of damaged copies.3. How reshuffling works
a. Reshuffling
Chromosomes.
Suppose one person has 2
identical copies of chromosome #1 and 2 identical copies of chromosome #2. (Draw these
chromosomes in one color, say pink.) Another person has 2 copies of chromosome #1 that are
the same as each other but different from the copies in the first person, and similarly
for chromosome #2. (Draw these chromosomes in another color, say white.) The offspring
of these two people will have a mixture of "pink" and "white
chromosomes. After several generations, it will be possible to get all conceivable
combinations of "pink" and "white" chromosomes.
(See problem 8-4 parts A & B.)
b. Reshuffling genes:
B. What happens to chromosomes during meiosis? -- see Handout 19B
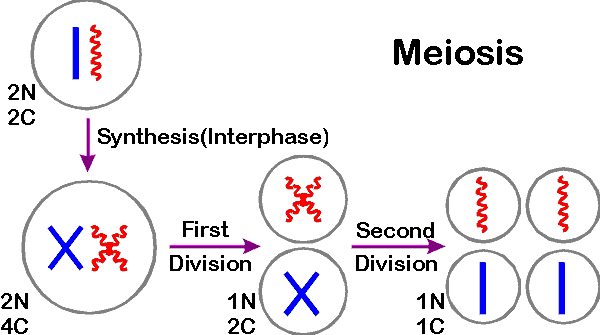
1. DNA synthesis occurs first -- before division. Meiosis is preceded by DNA duplication just as mitosis is. During the S before meiosis (or mitosis) the cell doubles the DNA content and # of chromatids per chromosome. So cell starts with pairs of doubled chromosomes = 4 copies of each chromosome.
2. Products: There are 4 products, each haploid (from meiosis), instead of 2 products, each diploid (from mitosis). To cut the number of copies of each chromosome from 4 to one requires 2 division, not one.
3. Two divisions of meiosis: The first division of meiosis separates homologs; the second division of meiosis separates sister chromatids.
4. What happens to N, c and # of chromatids/chromosome? The first division cuts the chromosome number per cell in half from 2N to N and cuts the DNA content per cell in half from 4c to 2c ("c" is defined below). The second division halves the DNA content per cell (from 2c to c), halves the number of chromatids/chromosome (from 2 to 1) and halves the total chromatid # per cell (from 2N to N). What happens in a cell with one pair of chromosomes is as follows:
Handout 19B summarizes c, N etc. for cells with one chromosome pair (N = 1) and for 3 pairs (N = 3). Handout shows chromosomes per cell at each stage (before S, after S, after 1st div. of meiosis and after 2nd div.)
See Becker fig. 20-3 (18-3) for a similar diagram of meiosis in a cell with 2 pairs of chromosomes.B. Definition of c
"c" is a measure of DNA content per cell, not the number of chromosomes or chromatids.
c = minimum DNA content per haploid cell of an organism = DNA content of haploid cell before S (with unreplicated chromosomes) = DNA content of one set of chromatids. C is NOT equal to N; c is the DNA content of N chromosomes (with one chromatid/chromosome).
To review Meiosis (so far), and compare to Mitosis, do or finish problems 8-1, 8-2 (parts A to E), 8-3, & 8-8 (parts A-D & G). Details of meiosis next time or see handout 19A.
II. The Mechanism of Meiosis -- see handout
19A.
A. Steps: These are diagrammed in detail on handout 19A, and comparisons to mitosis are summarized there. For similar diagrams of mitosis vs. meiosis see Purves 9.17 or Becker fig. 20-9 (18-9). For nicer pictures see Purves 9-14 or Becker 20-5 & 20-6 (18-5 & 18-6).
B. What if N >1? Handout 19A shows what will happen to a cell with 1 pair of chromosomes. (2N cell, N =1.) If there are additional chromosome pairs (N > 1), each pair will line up independently at metaphase I. This has important genetic implications, as will be discussed later.
C. Prophase I -- Some Differences from Mitosis
1. Crossing over. This is the time when recombination occurs by a "cut & splice" mechanism, which switches equivalent sections of chromosomes between 2 members of a pair. Recombination requires pairing, so homologous chromosomes are paired in pro. I of meiosis but not mitosis. More details to follow. (Pictures of pairing are shown in the texts.)
2. Duration. Prophase I in meiosis is generally much longer and more complex than prophase in mitosis. (Both texts divide prophase into early, mid and late substages; Becker 20-7 (18-7) has even more details if you are curious.) Pro. I can be very prolonged -- in human females, it lasts from before birth to the time the egg is shed. Consequences of this very long pro. I will be discussed next time.
D. Products of human meiosis (see Purves 43.3 (42.4) or Becker fig. 20-10 [18-10])
1. In Females: When female germ cells go through meiosis, the equivalent of 4 haploid nuclei are formed, but only one ends up in an egg. The genetic material that would end up in the "other 3" nuclei is shunted aside --it forms small structures called polar bodies. The egg contains (at least) the amount of cytoplasm that would be sufficient for 4 meiotic products and the genetic information of only one.
2. In males: When male germ cells go through meiosis, 4 sperm are formed.
III. Life Cycles
-- How do meiosis and mitosis fit
together? Or how does 1 multicellular organism give 2? Or better, for organisms
engaging in sexual reproduction, how do 2 (parents) give 1(offspring)?
A. Supercycle -- Handout 20B.
1. General idea -- Overview
Many different life cycles are possible, depending on the number of mitotic divisions that follow meiosis and/or fusion. The diagram on the top of handout 20 labeled "Supercycle" shows a generalized life cycle and explains the terminology. (Purves 9.12 or 29.2 is equivalent; Becker fig. 20-4 [18-4] is similar.) If an organism goes through all the stages shown, then it has both haploid and diploid phases.
2. Haploid vs Diploid Life Cycles
a. Diploid Life Cycle. What you get if meiosis → gametes. (No mitotic division of haploids).
b. Haploid Life Cycle. What you get if the zygote divides immediately by meiosis, not mitosis. (No mitotic division of diploids.)
3. Alternation of Generations
4. Super cycle can reduce to haploid or diploid life cycle
a. Diploid Life Cycle. If meiosis → gametes (no spores; no mitosis of haploids), the organism has really skipped most of the stages on the left half of the supercycle diagram and gone directly from germ cells to gametes. Such an organism has a basically diploid life cycle, as humans do. See Becker, fig. 20-4 (d) [18-4 (d)].
b. Haploid Life Cycle. An organism can skip most of the right half of the diagram if the zygote divides immediately by meiosis, not mitosis. (No mitosis of diploids. No germ cells.) Such an organism has a basically haploid life cycle. Some simple algae are like this. See Becker fig. 20-4 (b) [18-4 (b)].
4. Gametes and Spores -- Terminology
B. Some implications of this
cycle:
1. The Problem of Development. If zygote→ us by mitosis; how does development work? If all cells have the same DNA, why do cells make different proteins? In other words, why are the genes 'expressed' differently in different cell types? What sets and maintains the switches? How differential gene expression is set up and maintained is not entirely understood, but will be discussed some more next term.
2. Amniocentesis. If all cells of fetus have same genes/DNA/chromosomes, you can test the DNA or chromosomes of any fetal cell to look for abnormalities, even if the gene involved only affects (makes proteins in) certain specialized tissues. If you identify a fetus that will have sufficiently serious disabilities, you have the option to consider a therapeutic abortion. There are several current ways to test fetal cells, and more ways are under development. The most common current method is amniocentesis (testing fetal cells from amniotic fluid). Some examples: these will not be discussed in class but are included FYI.
a. Gene Mutations. You can look at DNA sequence (by PCR, use of probes, etc) for smaller changes affecting one or a few genes and/or nucleotides (so called "gene" mutations). Sometimes you can look at the protein the gene makes as in case (1) below, but sometimes you have to look at the DNA instead as in case (2). Some examples:
(1). Tay-Sachs disease. The gene that causes Tay-Sachs disease (when mutant) codes for an enzyme. The enzyme is made in many cell types, including the cells in the amniotic fluid. Using amniotic fluid cells, you can look at the DNA or measure the enzymatic activity of the protein made from the gene.
(2). PKU. The gene that causes phenylketonuria or PKU (when mutant) codes for an enzyme that is made only in liver cells. So using amniotic fluid cells, you can't measure the activity of the enzyme. But you can test the state of the gene itself (in amniotic fluid cells) to see if the gene is normal or mutant.
b. Chromosome Mutations. Since you can do banding, you can tell all the chromosomes and chromosome regions apart. Therefore you can detect large abnormalities affecting whole chromosomes and/or large blocks of genes (so called "chromosomal" mutations) from looking at karyotypes as discussed previously.
To review life cycles, cell cycle, etc. and how they all fit together, try 8-11 and/or 8-14.
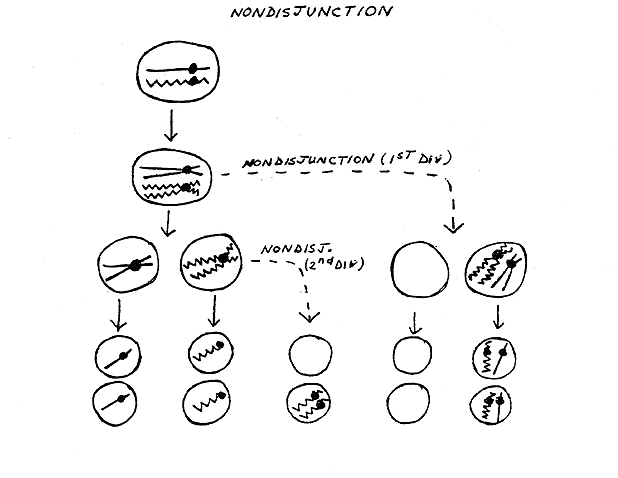
IV. Non disjunction -- see handout
20B bottom.
A. Where do individuals with missing and extra chromosomes come from?
Answer: Mistakes in Meiosis. Two types of mistakes:
Either way, nondisjunction gives gametes with extra and/or missing chromosomes (aneuploidy). See handout 20B, bottom half of page. When an aneuploid gamete (= gamete with missing or extra chromosomes) from one parent meets a normal gamete from another parent, then a monosomic or trisomic zygote is formed. The zygote can divide by mitosis to produce an aneuploid individual. Aneuploid zygotes containing missing or extra autosomes usually do not develop into viable individuals, but aneuploid zygotes containing missing or extra sex chromosomes (XO, XXY, XXX etc.) are usually viable as long as there is at least one X. (Why is this? See below.)
On handout 20B: Note that second division ND can involve either the "straight" or the "wiggly" chromatids, but only one case is shown. Also note that the "empty" cell is not really empty -- it is only missing a chromosome from the pair involved in ND. It has all the other chromosomes, but they are not shown to keep the picture as simple as possible. ND is an error than generally affects only one event at a time -- one pair of chromatids or one pair of homologs fails to separate at one stage of meiosis. usually all other separations of chromosomes and chromatids usually proceed normally. See Purves 9.18.
B. What types of aneuploidy are common? See lecture 19 for details.
1.
Trisomy 21.
2.
Aneuploidy of the sex chromosomes.
To review Nondisjunction, try 8- 8E & 8-9.
V. Inactive X's and Barr bodies
-- Why extra or missing X's are usually tolerated and extra or missing autosomes
are not
A. Lyon Hypothesis = inactive X Hypothesis
The idea that extra X's are genetically inert is called the inactive X or Lyon hypothesis. According to the Lyon hypothesis, every female is a mosaic, since some of her cells use her maternal X to make proteins and some use her paternal X.
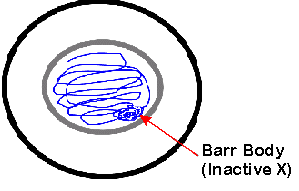
B. Barr bodies
You can actually see the inactive X during interphase because it forms a Barr body. There are 2 X chromosomes in every female cell, but (according to the inactive X hypothesis) only 1 of them works (is transcribed) most of the time. In general, if there are extra X chromosomes, all the extras are inactive, whether the cell is male or female. The inactive X's remain tightly coiled during interphase and are called Barr bodies. (So you can tell the sex of the cell without doing a karyotype.) Note that inactive X chromosomes are replicated, but not transcribed.
C. How is mosaic detected? Intro.
to Genetic Terminology
 Consider coat color in cats. This
is how Lyon actually figured out the inactive X existed. In cats, a gene controlling coat
color is on the X. The position of the gene is known as the locus
of the
gene. This gene has two alleles (alternate forms); one → black coat color and the other
→ orange. One of the alleles is present at the coat color locus on every
X. The Y chromosome
does not carry an allele of the coat color gene.
Consider coat color in cats. This
is how Lyon actually figured out the inactive X existed. In cats, a gene controlling coat
color is on the X. The position of the gene is known as the locus
of the
gene. This gene has two alleles (alternate forms); one → black coat color and the other
→ orange. One of the alleles is present at the coat color locus on every
X. The Y chromosome
does not carry an allele of the coat color gene.
Males have only one X, which carries either the black or the orange allele,
so normal male
cats are all black or all orange. (They may have regular stripes,
superimposed on the black or orange, but the background color is either all
black or all orange -- they don't have areas of orange and areas of black).
Females have two X's, so they carry two alleles of the coat
color gene -- one on each X. A female can be
homozygous black (have 2 black alleles), be homozygous orange (have 2 orange alleles), or
be heterozygous (have one allele of each color), as shown in figure. Females can be
orange, black or patchy (with areas of each color). Only heterozygous females
are patchy.
All this makes sense if only one copy
of the X works in each patch so only one copy of the coat color gene works per cell (and
per patch). Rare patchy males are XXY (Klinefelter's Kats).
Note "Patchy" is
called tortoise shell, not tabby; calico = patchy plus white. (Tabby = regular pattern of
stripes that occurs in both males and females.)
D. When do Barr bodies form? How do you get the mosaic?
Fertilized egg (zygote) →
ball of cells → each cell inactivates one X at random → each cell divides by
mitosis → descendants with same X on/off. Once an X is inactivated, it generally
remains inactivated through succeeding mitoses, so all mitotic descendants of a single
cell have the same X on and the same X off → all cells in one area (or with same
lineage) have same X on/off. Germ line cells (which will go through meiosis) turn both X's
back on before gametes are made (before meiosis occurs. So either one of the
two X chromosomes can be used or
inactivated in the next generation.
VI. Patterns of Inheritance -- An example
and the general principles -- See Top half of Handout 20A
A. What are the Big Issues to consider?
1. How are genes/genotypes inherited, and
2. How does a particular genotype (state of the genetic information) determine phenotype (appearance, function, etc)?
We'll start by looking more closely at the example of orange/black coat color in cats and then go on to other examples and the general case.
B. How do you figure out the pattern of Inheritance? -- an example for a gene on the X For a different classic example of inheritance of a sex-linked trait, see Purves 10.23 (10.25)
Note: The term "trait" is used in several different ways. It usually means whatever property you are following. Depending on the circumstances, it can mean the overall property you are considering such as coat color, OR it can mean the form (phenotype) of that property that you are following, such as orange coat color. So people speak of "the fur color trait" or "the orange color trait" depending on the context. ("Trait" is also sometimes used to refer to the carrier or heterozygous condition, as in "she has the sickle cell trait" meaning she has no symptoms but carries one allele for sickle cell.)
Consider a gene on the X such as the one we are talking about -- the one that determines orange vs black coat color. Suppose you mate a heterozygous female cat X orange male (see handout 20A). How do you figure out what will happen? Follow the steps below. (Each step is drawn on handout for each parent.)
1. Draw parental chromosomes with proper alleles.
2. Go through DNA replication to double DNA, chromatids/chromosome and # alleles/cell. Note sister chromatids are identical (if no crossing over**) but homologs need not be.
a. Sister chromatids must be identical since they are the 2 products of a single, semi-conservative, DNA replication. (See ** below.)
b. Homologs need not be identical -- one came from the mother and one the father. Homologs DO need to have the same genes (loci) lined up in the same order -- they just don't have to have the same alleles of these genes. In this case, one homolog has the orange allele of the coat color gene (at the coat color locus), and the other homolog has the black allele of the coat color gene.
3. Go through meiosis:
Homologs separate at first division and sister chromatids separate at second division.
This produces 4 gametes -- two different kinds, but in equal proportions (again assuming
no crossing over**).
**Note: Crossing over does not make any significant difference here because you
are following only one gene at a time. When you start considering two or more
genes at a time, then you have to take crossing over into account, and we'll
explain how to do that later. We're ignoring it now, because the gametes come
out the same (for the one gene under consideration) whether there is
crossing over or not. See Becker fig 20-14 [18-14] or Purves 10.19 (10.21). Crossing over
occurs in both figures, but you still get two gametes with one allele of the
gene (Y in Becker or B in Purves) and two gametes with the other allele (y or
b).
4. Do fusion of gametes from both parents to get zygotes (cats). You can use a Punnett square (or simple probability) to keep track of all combinations and proportions. This gives you the genotypes of the offspring (zygotes).
5. Look at genotypes and infer phenotypes of offspring = what develops from zygotes. Consider all possible combos and what proportions they occur in. In this case there is no dominance, so phenotype follows directly from genotype. If cat has only black (or only orange) alleles, you get a black (or orange) cat. If cat has both alleles, you get a tortoiseshell cat. (Cat is black in areas where X with B allele is on active X; cat is orange in areas where X with O allele is on active X. See below for mechanism of determination of black vs. orange.)
6. Terminology: In this course, and in many other contexts, the terms 'sex-linked' and 'X-linked' are used interchangeably, so sex linked = 'on the X chromosome.' If a gene is on the Y chromosome, a different term is used. Some biologists use the term 'sex-linked' to refer to genes on either the X or the Y. However, since there are very few genes on the Y, genes referred to as 'sex-linked' are almost always on the X (no matter how the term 'sex-linked' is used).
C. How does genotype determine phenotype?
How does a gene specify orange or black? In all cases, to figure out how genotype and phenotype correspond, you need to consider the enzymes and pathways involved. Current understanding of this case is as follows:
1. Black Part -- how is black pigment made?
| Gene A | Gene C | |||
| ↓ |
↓ |
|||
|
|
Enzyme A |
Enzyme C | ||
|
↓ |
|
↓ |
||
| Colorless Stuff | → | Black Stuff | → | Orange Stuff |
Gene A is probably on an autosome.
2. Orange part -- how is orange pigment made?
A gene on the X (call it gene C) codes for an enzyme that converts the black stuff into orange stuff. (The difference in color is probably caused by a different arrangement of pigment granules.) This gene has two alleles, called "O" and "B" above.
3. What determines orange vs black?
What differs between orange and black cats is the activity of the second enzyme. If the second enzyme is active, the black pigment is converted to orange. If the second enzyme is inactive, the black pigment remains black.
How the alleles of gene C determine color:
4. General Case.
For a sample problem on sex linked inheritance, try problem 9-9 A & C. (Part B depends on a discussion of dominance, which will be considered later.)
© Copyright 2006 Deborah Mowshowitz and Lawrence Chasin Department of Biological Sciences Columbia University New York, NY.