C2006/F2402 '04 -- Lecture 1 -- Updated 01/19/04
© 2004 Deborah Mowshowitz, Department of Biological Sciences, Columbia University, New York NY
Handouts: 1A -- Eukaryotic Cell Structure ; 1B -- Cytoskeleton; 1C -- Overview of Course (Brief Description)
Optional reading:References to Becker are to 5th ed. (When fig. numbers or pages in 4th edition are different, refs. to 4th ed. follow in parentheses.) Refs to Purves are to 6th ed.
Notes for this term will be extensive outlines, not word-for-word. This is partially for practical reasons and partially for educational reasons -- I think it encourages better student note taking and understanding. The outlines will be provided the night before (if not earlier) so you can print them out in advance and annotate them in class.
I. The Story So Far -- Summary of last Term
A. The Major Question: How do Living Things Work?
B. Answer so far has concentrated on
1. Prokaryotes (mostly) -- 1 intracellular compartment, unicellular
2. Macromolecular level -- did not discuss larger structures such as organelles, chromosomes, etc.
3. Looked at "big 5" issues = structure, function, manufacture (including energy requirements), regulation, and (evolutionary) origin
C. Bottom Line: DNA --> RNA
(& more DNA); RNA -->
protein --> job, say catalyzing X --> Y. Protein production & activity
are
regulated.
For recent developments on extensions of the central dogma see Scientific
American, Nov 2003 p. 46 "The Unseen Genome: Gems among the Junk," by Wayt
Gibbs. (A companion article by the same author is in the Dec. issue and
discusses chromosome structure and modification, which will be discussed later
this term.) To download the article, go to
Scientific American, and put the
words "unseen genome" in the
search box to get the link to the article. You may have to be at a Columbia
computer for this to work.
II. Overview of this Term
A. Similarities: Same Major Question; Same "big-5" issues.
B. Differences: Will concentrate on Eukaryotes (mostly); will look at the two big innovations of eukaryotes. -- Intracellular Compartmentalization & Multicellularity -- and the complications that result. See Becker table 4-1 for comparison of Eukaryotic and Prokaryotic cells.
C. Major Subjects of Term
1. Cell Bio -- study of consequences/implications of compartmentalization within cells (Lectures 1-9)
a. Macromolecules and bigger things (membranes and organelles -- chromosomes, etc.)
b. Co-ordination (between different parts of cell) -- how do things get to the right compartment? How do they cross membranes?
c. Regulation of eukaryotic gene transcription & the cell cycle -- how do things get made at the right time (as well as in the right place)?
d. Note: in this part, we will emphasize generalized features of all eukaryotic cells but discuss some specialized cell types.
2. Signaling & Cell-Cell communication
a. How cells communicate chemically (Lectures 10-13); how aberrations in communication may cause cancer.
b. How cell communicate electrically (Lectures 16-18 by Dr. Stuart Firestein)
3. Physiology
-- study of consequences/implications of multicellularity for maintenance
of homeostasis, movement & defense.
How specialized cells in a multicellular organism work to maintain a relatively
constant internal environment and how their
functions are co-ordinated. This subject will be divided as follows:
a. How the various systems (nervous, muscular, respiratory, etc.) work to maintain homeostasis of temperature, gas, fluid, salt & water (Lectures 14, 15 & 19-21
b. Immunology -- The most specialized system; how a multicellular organism fends off invaders (Lectures 22-23)
4. Development -- How to build a multicellular organism. How cells get to the right place in a 3D organism; how the cells specialize and how they stay that way. Lectures 24-25.
III. Eukaryotes -- a closer look.
A. What can you see in the light microscope? Suppose we draw a standard light microscope view of eukaryotic cell -- what can you see? Can't actually see membranes or details of many organelles.B. Issues of resolution (For pictures using the different methods, see Becker fig. 1-3 or Purves 4.3.) For more details on microscopy, see Guide to Microscopy (a supplement included with the 5th ed.) or appendix (4th ed.)
Types of Microscopes -- Light microscope, SEM (scanning electron microscope) and TEM (transmission electron microscope)
|
Limit of Resolution |
Useful Magnification |
Features |
|
|
Eye |
0.1 mm |
1 |
|
|
Light Microscope |
0.2 micron |
1000X |
Can look at living specimens |
|
TEM |
1-2 nm (in practice) |
100,000X or 100X light scope |
Detect electrons from source that pass through thin slice of tissue |
|
SEM |
Larger than TEM but gives 3D picture of surface |
Less than TEM |
Detect secondary electrons emitted from surface of sample |
Sizes of things commonly encountered in biology
|
1/1000 |
1/1000 |
1/1000 |
1/10 |
|||||
|
m (meter) |
---> |
mm |
----> |
micron |
---> |
nm (nano) |
----> |
Angstrom |
Sizes of some cells, structures and very small living (?) things:
| Cell Type | Diameter |
| Typical eukaryotic animal cell | 50 microns (mu) = 50,000 nm. |
| Typical bacterium | 1-5 microns = 1-5000 nm |
| Smallest known bacterium | 0.2 microns = 200 nm |
| nanobe (for Times article see; for more info see) | 20-150 nm |
| ribosome | 50-60 nm |
For more examples and tables of relative sizes see Becker fig.1-2 & box 1A or Purves 4.2 .
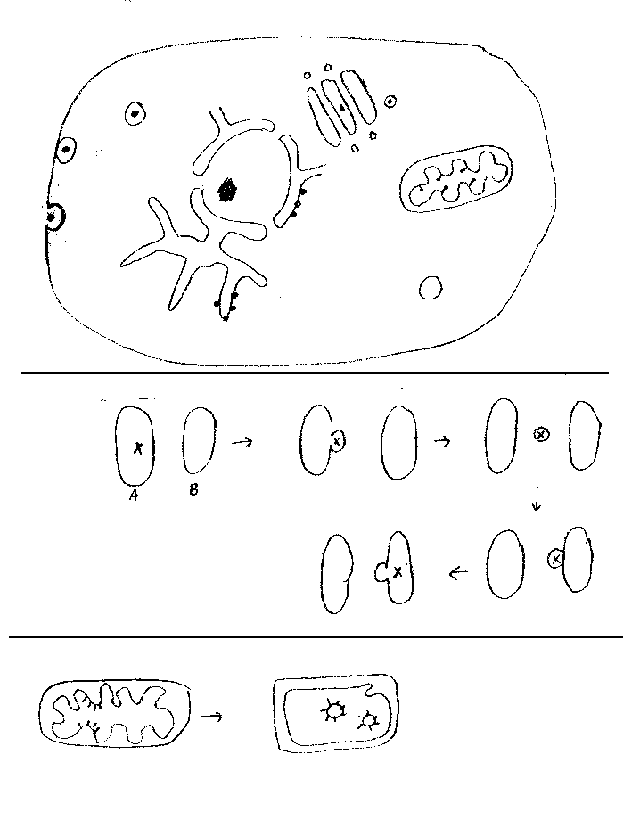
C. What you can see in the EM? See handout 1A and fig. 4-5 of Becker or 4.7 of Purves for pictures of whole cells. Some specific references to pictures of organelles are given below, but all pictures in Ch. 4 of both texts are well worth it. In the EM you can actually see membranes; structure of membranes will be covered next time. What else can you see in a typical animal cell?
1. Nuclear details (See figs. 4-4 & 4-10 of Becker or 4.8 of Purves)
Nucleolus -- not membrane bounded
Nuclear Pores & Nuclear Envelope (Double membrane with holes punched in it)
Perinuclear Space (Space between two layers of nuclear envelope)
No ribosomes in the nucleus.
2. Endomembrane system (See Purves figs. 4.11 to 4.13 or Becker fig. 4-15 to 4-18)
Components
a. ER (smooth & rough) -- continuous, but only rough (RER) has attached ribosomes
b. Golgi
c. vesicles (lysosomes, transport vesicles, secretory vesicles, endosomes).
Function of components
a. Rough ER, Golgi, most vesicles: Involved in processing, sorting, packaging and transporting proteins to their proper destination -- to right part of EMS or outside the cell.
b. Lysosomes: contain acid hydrolases -- used to degrade/recycle macromolecules.
c. Smooth ER: lipid synthesis, Ca++ storage, detoxification, etc.
Isolation: See methods section below for how the various components are isolated and/or distinguished from each other.
Structure of Components
Membranes of ER continuous with outer layer of nuclear envelope
All parts are made of vesicles and flattened sacs (cisternae)
Each vesicle or sac surrounded by a single membrane (one bilayer)
Inside of each vesicle/sac = lumen = space inside a hollow organelle, organ, or tube.
Exocytosis and endocytosis-- see handout 1A
All internal spaces of endomembrane system (lumens & perinuclear space, but not inside of nucleus) are effectively connected to each other and the outside of the cell
How are these spaces & outside of cell (effectively) connected? Through formation and fusion of vesicles.
Vesicular traffic (budding off and fusion of vesicles) moves and carries "cargo" and membranes from one membrane-bound compartment to another.
Vesicle traffic can transport cargo two ways (see handout 1A).
(a) from one compartment inside cell to another or
(b) between inside and outside of cell (by exo- & endocytosis).
You need labeling of cargo to determine direction.
3. Peroxisomes (See Becker fig. 4-19 & 4-20 or Purves 4.19)
Surrounded by a single membrane (like lysosomes; unlike nucleus or endosymbionts).
Enzyme content -- lumen contains many oxidases (in animal cells for detoxification, breakdown of long chain fatty acids) and catalase (for destruction of hydrogen peroxide produced by oxidases).
Contain enzymes producing dangerous (highly reactive) products such as H2O2. (Note that the enzymes themselves are not potentially dangerous -- the enzymes do not catalyze breakdown of normal macromolecules. It's the products of enzyme action, such as peroxide, that are dangerous.)
Not part of endomembrane system -- do not bud off from Golgi.
How are these distinguished from lysosomes? See methods below.
4. Location/function of ribosomes.
Ribos are in cytoplasm either "free" or attached to ER.
Free ribos make proteins for nucleus, cytoplasm, peroxisomes etc. -- all parts of cell except EMS.
Bound ribos on RER make proteins for EMS or outside the cell.
5. Mitochondria and chloroplasts
Have own genetic systems (DNA, ribosomes etc. -- everything needed for DNA replication, transcription & translation.)
Most proteins of mito and chloro are NOT made inside the organelle -- most are encoded in the nucleus, made in the cytoplasm, and transported into the organelle after synthesis.
There are strong similarities between mito &. chloro -- electron transport in membrane used to pump protons, H+ gradient used to make ATP; carbon rearrangements carried out in matrix or equivalent, etc. Exergonic process that drives electron transport is different -- light absorption (in chloro) vs oxidation of reduced carbon cmpds (in mito).
Origins -- probably endosymbionts. Mito. and chloro. were once probably free living bacteria. (See Purves 4.15 or Becker Box 15A)
Q's to ask yourself: (1) What are the major similarities and differences between peroxisomes, lysosomes and mitochondria? How can you tell them apart? It is a good idea to make a table that compares and contrasts the three organelles. (2) How are new organelles made? The answers are different for the 3 types of organelles and will be discussed in a few lectures.
5. Cytoskeleton (see handout 1B & Becker figs. 4-23 & 4-24 or Purves 4.21) -- we may not get to this until next time.
Structure & properties of each type are summarized in table on 1B (see texts for pictures)
Functions
a. Support/strength -- weight bearing, shape determining.
b. Movement (MF & MT)-- most movement involving MF & MT occurs by sliding of one fiber or organelle relative to another. (Changes in length due to polymerization and depolymerization do occur.)
c. Localization of other factors -- act as peg board or framework for attachment of organelles, enzymes, etc.
IV. Methods -- these will be discussed in Lecture #1 or #2 as we get to them.
How do you find out which components are in each cell part?
Reminder:
Becker has a guide to all techniques and methods described in the book -- see
inside front cover in 5th ed. (pp. xiii -xv in 4th).
A. "Grind and Find" (Biochemical separations and assays). See Becker Box 12A pp. 326-330 (332-336).
1. Grind: Break up cell into parts, and separate (fractionate) parts by ultracentrifugation.
2. Find: Assay (test) each cell fraction for enzymes of interest. Find the cell part associated with each particular enzyme.
B. In situ labeling (Localization of enzymes "in situ" = in place)
1. Substrate: Enzyme substrate is soluble, so it diffuses to site of enzyme.
2. Product: Product of enzyme catalyzed reaction is insoluble, so it stays in the place where it is made..
3. Localization: Insoluble product is produced and precipitates only at location of enzyme -- pin points position of enzyme
4. Detection of product: Can be colored (for detection in light microscope) or electron dense (for detection in EM). For an example, see Becker fig. 12-20.
C. Immunofluorescence -- Use of labeled antibodies as tags -- direct (one step) and indirect (two step). See Becker Guide to Microscopy pp 8-11 (Appendix, pp. 824-827) and table 22-2. For a picture of a typical result, see Becker 22-1 or Purves 4.3. A handout detailing the procedure will be provided next time. Allows you to visualize location(s) of particular proteins.
Next time: More on the cytoskeleton, and the structure of cell membranes.
{kind=link}