C2006/F2402 '04 OUTLINE OF LECTURE #10
(c) 2004 Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last update 02/24/2004 08:52 AM . There was an error in earlier version of outline -- see note on III. A. (highlighted in blue).
Handouts: 10A-C. 10 A -- Regulation of intracellular Fe levels + cell cycle info (not on web). 10B-- Alternative splicing. 10C -- Overview of Signaling -- Big Bang model + Types of signaling molecules (not on web). Extra copies of all handouts are in boxes on 7th floor of Fairchild.
I. Details of transcription in eukaryotes (as vs. prokaryotes), cont.
A. Regulatory Sites in the DNA -- see last lecture, and handout 9.
B. Regulatory proteins or TF's = transcription factors
1. Basal TF's. Needed to start transcription in all cells. (See Purves fig. 14.14 (14.16) or Becker fig. 19-14.)
a. Many basal TF's needed.
b. People usually are interested in basal TF's for RNA pol. II. (since pol II --> mRNA)
(1). These are called TFIIA, TFIIB, etc.
(2). Major one is TFIID; it itself has many subunits. Most studied subunit is TBP (TATA binding protein -- See Becker fig. 19-15.) Recognizes TATA box when there is one.
c. Basal TF's bind first to core promotor, and then RNA pol binds to them. Takes a lot of proteins to get started. RNA polymerase does not bind directly to the DNA.
2. Regulatory or Tissue Specific TF's -- used only in certain cell types or at certain times.
a. Bind to areas outside the core promotor -- usually to enhancers or silencers (distal control elements) but sometimes to proximal control elements
b. When regulatory TF's bind, can decrease or promote transcription.
(1). Activators. TF's called activators if bind to enhancers and/or increase transcription.
(2). Repressors. TF's called repressors if bind to silencers and/or decrease transcription.
(3). Co-activators. Proteins that connect TF's to each other (but don't bind directly to the DNA) are often called co-activator (or co-repressor) proteins.
c. Co-ordinate control. A group of genes can all be turned on of off at once in response to the same signal (heat shock, hormone, etc.). These genes do not need to be near each other -- they just have to have the same control elements. See Purves 14.16.
(1). Common control elements: All genes turned on in the same cell type and/or under the same conditions have the same control elements -- therefore these genes all respond to the same TF's. Result is multiple mRNA's, all made in response to same signal (s).
(2). Compare to situation in prokaryotes:
|
|
Prokaryotes | Eukaryotes |
|
Co-ordinately controlled genes are |
Linked |
Unlinked |
|
mRNA is |
polycistronic (1 mRNA/operon) |
moncistronic (1 mRNA/gene) |
|
Operons? |
yes |
no |
|
Control elements are found |
once per operon |
once per gene |
| Control can be positive or negative but is more often | negative --repressors needed to turn gene off | positive -- activators needed to turn gene on. |
d. Structure & Function of regulatory TF's is modular
(1). Each TF has multiple domains.
(a). Each TF has a DNA-binding domain -- specific for particular sequence(s) and/or gene(s)
(b). Each TF has a transcription regulation domain (also called trans acting domain or in many cases transcription activating domain) -- determines effects of DNA binding by given TF (activation vs inhibition of transcription)
(c). TF's that are hormone receptors also have a hormone-binding domain.
(d). TF may have additional domains, such as dimerization domain. Many TF's must dimerize to work. (Monomer is inactive.) Some form dimers with other molecules of the same protein (result is a homodimer) and some form dimers with a different protein (result is a heterodimer).
(2). Modules (domains) can be switched -- Recombinant DNA methods can be used to make hybrid TF's. This has many uses in research; some examples are in the problem sets.
(3). How do regulatory TF's act?
By binding to control elements with their DNA-binding domains and to basal TF's or each other (or to co-activator or co-repressor proteins) with their transcriptional regulation domains.(4). Types of DNA-binding domains. These are often classified by the shape of the DNA binding region. Some common DNA binding shapes (called structural motifs) are listed below for reference only (FYI). For pictures, see Purves p. 273 (6th ed) or fig. 14.18 (5th ed) or Becker fig. 21-25.
Zinc finger
leucine zipper
helix-loop-helix
helix-turn-helix.
To review transcription, try problems 4-16 and 4-17 A.
II. Regulation at Splicing -- Alternative Splicing
A. There are two ways to get a collection of similar proteins
1. Gene families -- multiple, similar genes exist due to duplication and divergence of genes. Example: the globin genes constitute a family. Different family members code for myoglobin, beta-chains, alpha-chains, delta-chains, etc.
2. Alternative splicing etc (See C below) -- only one gene, but primary transcript spliced in more than one way.
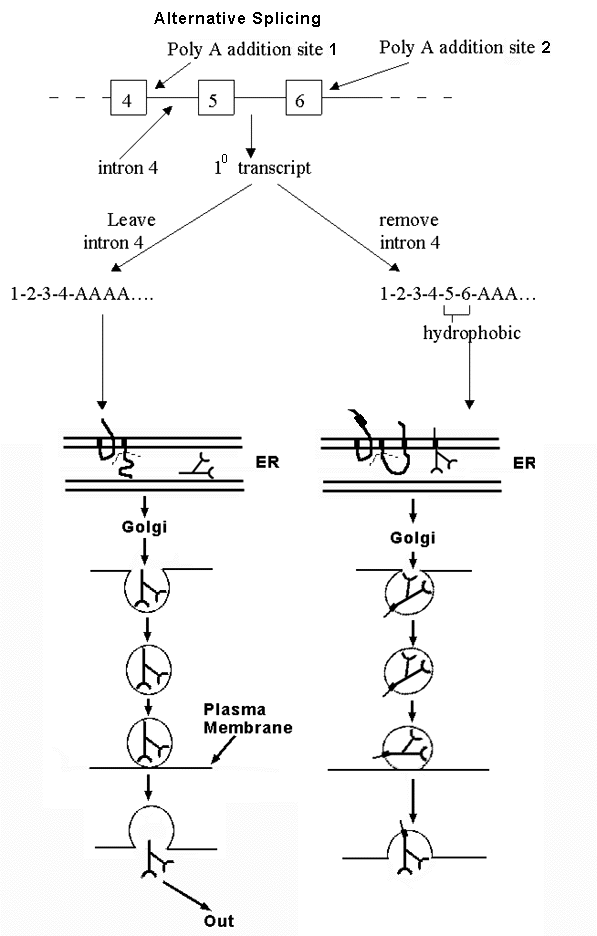
B. An example of alternative splicing -- see handout 10 B and Becker fig. 21-31 -- how to get either soluble or membrane-bound antibody from alternative splicing of the same transcript.
1. Antibody can be membrane bound or secreted. Fate of antibody depends on whether peptide has a stop transfer sequence or not.
a. If has stop transfer, locks into membrane of ER and goes thru Golgi etc. to plasma membrane; stays in membrane.
b. If has no stop transfer, enters lumen of ER, goes thru Golgi etc. and then is secreted.
2. Gene has two alternative polyA addition sites. Which one is used determines final location of protein.
a. If one (at end of exon 4) is used, protein contains no hydrophobic stop transfer sequence, and protein is secreted.
b. If other one (at end of exon 6) is used, protein contains hydrophobic sequence encoded by exons 5 & 6, and protein stays in plasma membrane.
3. mRNA can be spliced and/or poly A added in two alternate ways. Location of protein (antibody) depends on whether splicing of intron 4 or poly A addition happens first. Think of it as a competition. Either
a. Poly A adding enzymes get there before the spliceosome. In that case, poly A is added to site near end of exon 4, and rest of intron 4 (and rest of gene) is never transcribed, or
b. The spliceosome gets there first. In that case, Intron 4 is transcribed and spliced out before poly A can be added. (In this case, poly A is added at the end of exon 6 instead.)
4. Why are 2 forms of antibody needed?
a. Membrane-bound form of antibody serves as receptor for antigen = trap to detect when antigen is present. Binding of antigen (ligand) to antibody (receptor) serves as trigger to start secreting antibody.
b. Secreted form binds to soluble antigen and triggers its destruction in multiple ways.
C. The general Principle -- You can get many different proteins from a single gene by the processes listed below. Therefore biologists are interested in proteomics (study of all proteins made in a cell) not only genomics (study of entire DNA or gene content).
1. Starting transcription at different points
2. Ending transcription (adding poly A) at different points
3. Splicing out different sections (exons as well as introns) of the primary transcript -- alternative splicing.
To review regulation so far, try problems 4-9 to 4-11. To review the details of alternative splicing, try problem 4-12.
III. Regulation at translation.
A. How to control rate of translation? In principle:
1. Can regulate half life of mRNA (control rate of degradation). In prokaryotes most mRNA's have a short 1/2 life; in eukaryotes this is not necessarily so. Different mRNA's have very different half lives. (Note: proteasomes degrade only proteins NOT RNA's. )
2. Can regulate rate of initiation of translation (control how effectively translation starts)
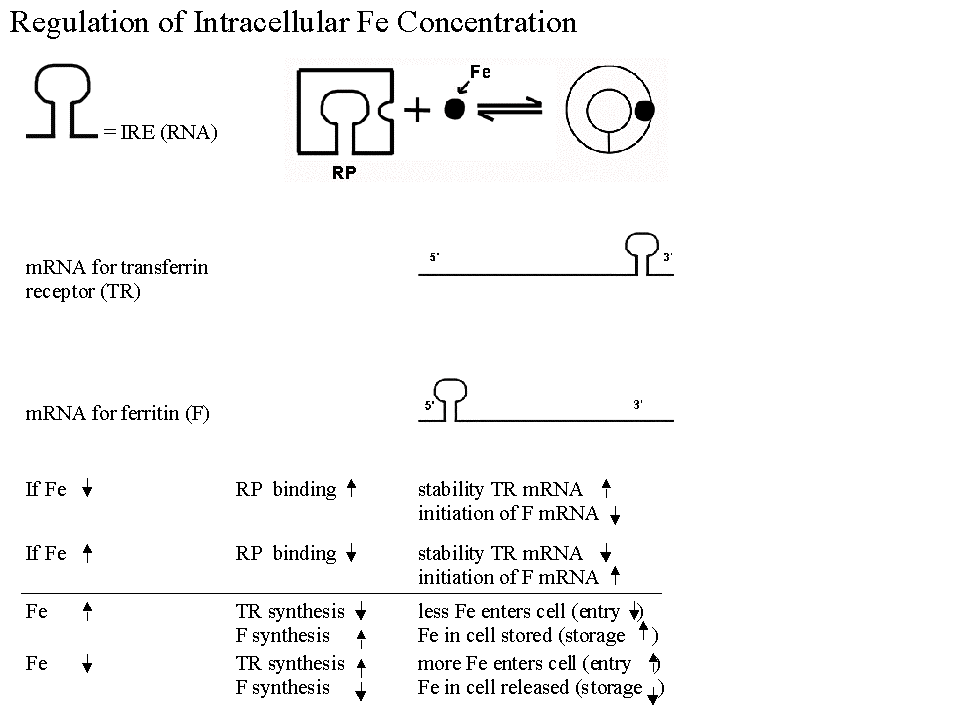
B. Most Famous Example of Regulation of Translation. Regulation of synthesis of Ferritin & Transferrin Receptor (& intracellular iron levels). See handout 10A & figs. 21-33 & 21-34 in Becker.
1. In best known example, have a regulatory protein with following properties:
a. Protein controls half-life of one message by binding to sequence at 3' end and blocking degradation.
b. Protein controls usage of a different message by binding to sequence at 5' end and blocking initiation of translation.
c. Protein is allosteric and has two forms -- binding of a small molecule effector (Fe in this case) determines which form protein is in, and therefore what effect(s) it will have.
d. System is similar in many respects to induction/repression but regulator protein acts on a sequence in the mRNA (called an IRE - iron response element) instead of on DNA. Comparison to induction/repression:
|
Similarities |
Differences |
|
IRE = cis-acting site like an operator |
This RP binds to sequence on mRNA NOT to DNA |
|
RP = trans-acting regulatory protein like a repressor = allosteric protein with an allosteric effector (Fe) |
Binding of RP affects translation, NOT transcription. |
2. What are the important features of this system for cell function?
a. Works to stabilize intracellular iron levels
b. System is self-correcting -- Deviations in either direction (if iron is too high or too low) are corrected back to standard.
c. There are two opposing correcting factors or processes, not just one. (There are mRNA's for two proteins -- ferritin & transferrin receptor, and the two proteins have opposing effects, either raising or lowering intracellular iron levels.)
3. Negative Feed Back. A self-correcting system like this one is called a negative feedback system. More examples (& features) of negative feedback will be discussed when we get to homeostasis.
IV. Post Translational Regulation. Don't forget: regulation occurs after translation too -- after proteins are made, they can be modified. Many examples of post translational modification have already come up and more will be discussed below.
A. Proteins can be modified covalently either reversibly (for ex. by phosphorylation and dephosphorylation), or permanently (for ex. by removal of N-terminal met., addition of sugars -- glycosylation, etc.)
B. Proteins can be activated or inhibited by reversible noncovalent binding of other factors -- small molecule allosteric effectors, other proteins such as calmodulin, etc.
C. Proteins can be selectively destroyed. Not all proteins have the same half life. Major factor in regulation of protein turn over is control of addition of ubiquitin leading to destruction by proteasome.
To review post-transcriptional &/or post-translational regulation, try problem 4-13.
V. Regulation of the Cell cycle -- Regulation of more complex event/process. We've looked at how to regulate one protein or a few at a time. Now how to regulate the cell's main event? Involves many steps, utilizing many of the different types of regulation discussed above. See handout 10A.
A. Review of G-1, S etc.; histones made in S too along with DNA.
B. Mutants indicate the existence of two major control points or checkpoints. See Becker fig. 17-30 or Purves 9.6.
1. Near G1/S (border of G-1 & S) also called "start" in yeast or the "restriction point" in mammalian cells
a. Actual checkpoint is in G-1, near end; determines whether cell will enter G-0 (non dividing or non cycling state) or S (or pause in G-1 and await further instructions). Irreversible decision is to proceed past checkpoint and enter S.
b. This is the major checkpoint/decision point for animal cells in the adult.
c. Many growth factors (like EGF) act at this point -- are needed to enter S.
2. G2/M (Border of G-2 & M).
a. Needed to check all components are ready for mitosis before proceeding.
b. This is the major checkpoint/decision point for cleaving eggs. Cycle here is basically all S and M -- G1/S switch is in override.
c. Note: Passage of this checkpoint in animal cells generally depends on the internal state of the cells, not on presence/absence of GF's.
3. Additional checkpoints exist; see texts (especially more advanced ones) if you are interested.
C. Basic switch or regulatory protein -- similar in both cases:
1. Positive or negative control? Fusions (between cells at different points in the cell cycle) imply decisions at checkpoints are controlled primarily by an "on switch" not release of an "off switch." See Becker Fig. 17-31. You need presence of an activator signal (not loss of an inhibitor) to proceed past a checkpoint. (However, production/activation of the switch protein complex involves a complicated process which can involve multiple inhibitors and/or activators.)
2. Switch controlled by, or is, a protein kinase -- at least 2 different ones -- one for "start" and one for G2/M. In each case you need to have an active protein kinase.
3. Each kinase phosphorylates a specific set of proteins
| Kinase | Acts at | Phosphorylates | Effects |
| MPF | G2/M | histones, lamins, MAP's | Change State of Nucleus (Envelope & Chromatin) & Spindle |
| Start Kinase | G1/S | TF's or TF inhibitors (ex: Rb protein) | Trancribe Genes --> new proteins needed for DNA & histone synthesis |
4. These kinases have 2 parts
a. CDK = cyclin dependent kinase (called p34, cdc2 etc.). This is the actual catalytic protein. Level of kinase protein itself remains steady in cycling cells. (Probably degraded if cells stay in G0 = exit the cell cycle.) Inactive without cyclin; therefore catalytic activity of the kinase is dependent on cyclin (& other factors; see below).
b. Cyclin -- builds up, peaks, degraded (in proteasome), repeats (See Becker fig. 17-33 for graph). Acts as an activator of the CDK.
c. Complex of CDK + cyclin forms; inactive; action of right combo of (additional) kinases and phosphatases --> active form. (See Becker fig. 17-34 if you are curious about the details)
d. Different cyclins for G1/S and G2/M; usually different CDK's too (Depends on organism whether same or different).
5. Effects of kinase activation
a. Allow cells to pass checkpoint and enter next phase of cell cycle
b. Kinase phosphorylates and thus activates proteins needed to successfully complete next stage of cycle -- start DNA replication, disassemble nuclear membrane, separate chromosomes, etc. (see table above)
D. Overall Cycle as shown in texts (Probably an over-simplified view) -- activation of CDK triggers G1-->S and then CDK is inactivated by degredation of cyclin. Process repeats at G2-->M (with different cyclin/CDK complex). See Purves fig. 9.7 or Becker fig. 17-35 or 17-38 [17-37] or handout 10C.
| Modifications (kinases, phosphatases etc.)** | ||||||||||
| CDK (stable) | | | CDK (inactive) | ||||||||
| --> | Inactive Complex |
----------> |
Active Complex (Kinase) | --> | Add P to target proteins# | --> | Continue Cycle | --> | ||
| Cyclin (newly made)* | Cyclin Degraded | |||||||||
| | | ||||||||||
|
* Synthesis of cyclin triggered by: |
||||||||||
**Many "brake" proteins (for example, p53) act at this step
# See table above for target proteins
1. Why all these multiple controls and steps? If adult cell divides when it shouldn't --> cancer; if fails to divide get loss of repair (no healing) and degeneration in adult. So need this very carefully controlled. So have multiple "brake" and "accelerator" proteins in this system. Many cancers traced to loss of "brake protein" or over production/activation of "accelerator protein," as will be discussed in lecture 12.
2. Regulation occurs at many levels
a. Cyclin levels are regulated by controlling synthesis of mRNA (transcriptional control) and protein, and by controlling degradation of mRNA and protein (post transcriptional control)
b. Activity of many proteins involved is regulated post translationally by extensive modifications (mostly phosphorylations and dephosphorylations) -- examples include kinases and TF's.
c. Cycle is influenced by external factors (GF's, hormones, contact from other cells, etc.) and by internal factors such as state of the chromosomes, DNA damage etc. Both sets of factors alter passage through the cycle by triggering activations/inactivations of proteins (TF's, kinases, etc.). Regulation of cell cycle integrates both sets of information.
To review regulation of the cell cycle, try Problem 15-8.
IV. Introduction to Signaling -- How are messages sent from one cell to another? How are events in a multicellular organism co-ordinated?
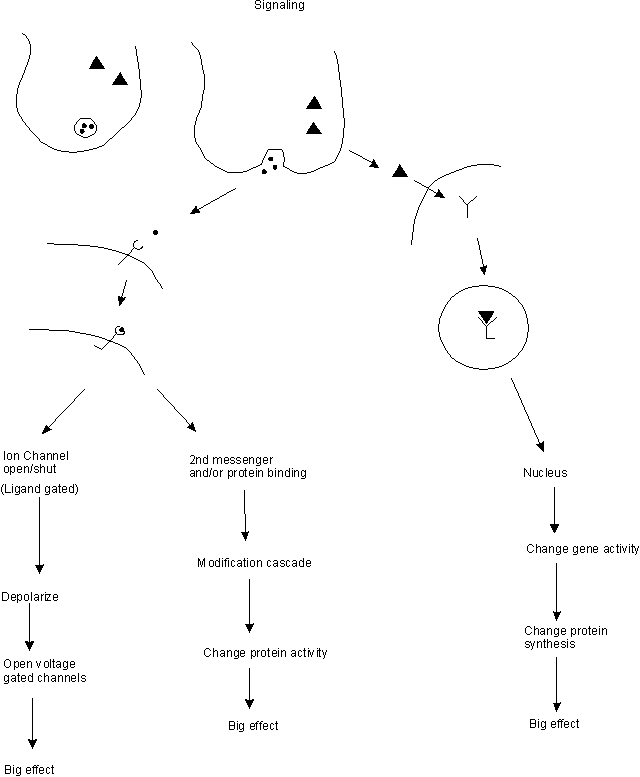
A. Usual Method -- one cell secretes a signal molecule that binds to a receptor on (or in) the target cell --> amplification --> big effect.
B. How do secreted signal molecules work at molecular level? Overview. See top part of handout 10C.
1. Signals are evolutionarily conserved -- same signal molecules used by different organisms for different purposes.
2. Big Bang Theory. All signals --> amplification = big effect from a small concentration of signal. How is amplification achieved?
a. By affecting transcription/translation --> lots of new protein --> big effect
b. By cascades of modification and/or second messengers (see below) --> lots of modified protein --> big effect
c. By opening (ligand-gated) channels --> ion flow --> open more (voltage-gated) channels --> big change in ion concentrations --> big effect
3. Types of Signals. Two main kinds of chemical signals -- lipid soluble and water soluble
| Signal Type | Example | Receptor Type | Effect |
| Lipid Soluble | Thyroxine, steroids | Intracellular | Gene activity |
| Water Soluble | Peptide hormones, GF's | Extracellular | Protein activity (usually) |
4. Types of Receptors -- intracellular and extracellular
a. Intracellular -- all members of same gene/protein family. All Transcription factors.
b. Extracellular -- three main types.
| Receptor Type | 2nd Messenger? | Example = Receptor for | Usually Affects | |||
| G protein Linked | Yes (cAMP, IP3 & DAG, or Ca++) | Epinephrine & many other hormones | Protein Activity | |||
| Tyrosine Kinase (TK) Linked | Sometimes (IP3) | GF's, insulin | Gene or Protein Activity | |||
| Ion Channel* | No | Acetyl Choline (nicotinic) | Ion flow |
* To be discussed at length when we get to nerves. The other two types will be discussed in lecture 11 or 12.
C. Major types of secreted Signals -- classified by type of cell that makes them and/or target location. See bottom of Handout 10C for pictures -- numbers of pictures match numbers below..
1. Endocrine: Endocrine gland secretes signal molecule (hormone) into blood ---> target (often far away)
2. Paracrine: Receptor is on adjacent cells. Act locally. Examples:
a. histamines (mediate allergic reactions, responses to inflammation)
b. prostaglandins -- initiate uterine cramps; cause fever in response to bacterial infection.
3. Autocrine: Receptor is on same cell. ex. = some growth factors
4. Neurocrine: Neuron secretes neurotransmitter, acts on receptors on neighbor (gland, another neuron or muscle) -- like a paracrine.
5. Neuroendocrine: Neuron secretes signal molecule that acts like a hormone (travels through blood to target).
6. Exocrine: Exocrine gland secretions (exocrine = released outside the body) can carry signals --> target in different individual = pheromones (detected by olfactory receptors in mammals).
D. Other types of Signaling
1. Gap Junctions -- allow ions & currents to flow directly from cell to cell -- used in smooth muscle --> synchronized contractions.
2. Juxtacrine. Cell surface proteins from two different cells contact -- used in immune system. Similar to basic system, but signal molecule is not secreted -- remains on cell surface.
Next time: How do signal molecules work to get a response in target cells?
{kind=link}
{kind=link}
{kind=link}