C2006/F2402 '04 -- Outline for Lecture #25 -- Molecular Aspects of Immunology
(c) 2004 Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last Update: 04/28/04 12:43 PM .
Handouts: See 23B for order of class switching.
25A -- Structure
of Antibodies and their respective genes (posted version is labeled 24D)
25B -- Molecular
basis of Antibody production and class switching in 1o & 2o
response. (posted version is an older, hand written version, labeled 24B)
I. How do T and B cells get activated? Wrap Up. See handout 24B (picture on bottom) & 25C (table).
See Recitation Problems #12, Questions 1 & 2. (RP #12 will be posted on line shortly.)
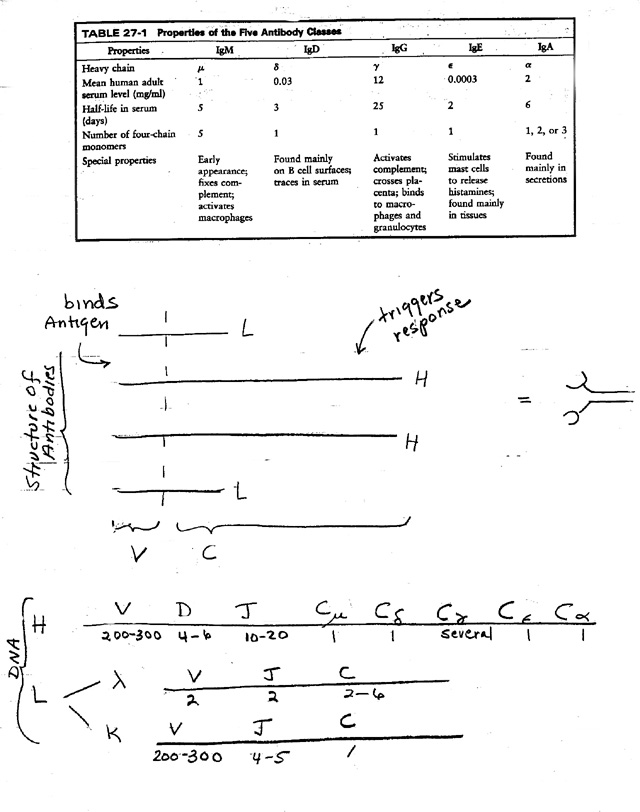
II. Ab Structure -- How is specificity achieved? (Handout 25A -- middle or Purves 19.11)
A. V vs C -- types of Immunoglobulin (Ig).
1. There are 5 main classes of Antibody -- IgM, IgD, IgG, IgE, and IgA. See table on top of handout 25A & Purves Table 19.2.
2. V & C: Each Ab or Ig is made up of a V section ("variable" region) & a C section ("constant" region).
3. Variable region
a. V is specific for Ag. Determines what Ag will be bound = grabbers.
b. V is variable due to differences in sequence, not just differences in folding around Ag.
c. Every Ab or Immunogloblin (Ig) has (at least) 2 grabbers.
d. All grabbers in one Ab are the same.
e. All the antibodies made by one Ab-producing cell have the same V. All the antibodies made by descendents of that cell have very similar V's. (Minor differences are due to somatic mutation; see below.)
4. Constant region
a. C determines biological effects -- localization of Ab, and what will happen as consequence of binding Ag. (Whether complement will be activated, whether Ab will be found primarily in blood or secretions, etc.)
b. 5 main types of C regions, therefore 5 main classes of antibody. (For properties of the dif. classes see handout 25A top or Purves Table 19.2)
c. The same V's can go with different C's. (Called "Class Switching")
(1). All the antibodies made by one Ab-producing cell do not necessarily have the same C.
(2).The antibodies made by descendents of a single cell may have different C's. The same variable region can go with different constant regions as B cell clone expands. How is this possible? Need a closer look at Ig structure
Try Problems 13-1 & 13-2.
B. H vs L. See handout 25A middle or Purves 19.11
1. Every Ig has 2 kinds of chains, L ("light") and H ("heavy"). Light and heavy refer to relative differences in mol. wt.
2. Basic unit is 2 of each for a total of 4 chains. (For number of basic four-chain units per Ig, see table.)
3. Variable region (grabber) made of parts of each.
4. Each chain has a constant region
a. 2 kinds for L (kappa or lambda)
b. 5 basic kinds for H (mu, delta, gamma, epsilon or alpha)
c. Hc (constant part of H) determines class (IgM, IgD, IgG, IgE, IgA)
d. Class (determined by Hc) determines location & other aspects of function (see "special properties" in table)
d. Class switching involves the H chains only, not the L chains.
5. Myelomas & Hybridomas: Ig structure was figured out by studying proteins made by myeloma cells (cancers derived from Ab-producing cells) or hybridomas (hybrids of Ab-producing normal cells and cancer cells). Only way to get large numbers of cells all making the same Ab/Ig. See texts for significance of hybridomas and monoclonal antibodies. (Purves 19.13)
Try problem 13-3.
C. Classes of Ab and class switching during development of immune response
1. Order of events (will be on class handout, not on web) during immune response
a. First make M, then M + D -- all on surface.
b. Meet Ag --> primary response: secrete M.
c. Meet Ag a second time --> secondary response; secretes usually G but can be E or A.
d. All these Ig's combine with same Ag
2. Implications of structure and switching
a. Can make different variable regions -- zillions of them, one for each dif. epitope. So the IgG, for example, in a person is a mixture -- all IgG molecules have the same constant regions but have different variable regions.
b. During differ. stages of immune response, can make Ab with same variable region but different constant region (for H). Can switch class and/or secreted vs. surface. How is this possible?
3. What we already know: How switch from membrane bound to secreted works. Variable part stays same; Hc changes from hydrophobic to hydrophilic by alt. splicing/poly A addition.
4. What we don't know so far: How do you make so many dif. variable regions AND What changes when you switch classes (from IgM to IgG? M to M + D)? Is it alt. splicing? Must be rearrangement at DNA or RNA level. Which is it?
III. Structure of the DNA -- Basis of Class switching and generation of diversity (G.O.D) -- handout 25A, bottom & 25B
A. Basic idea: genes for H and L are mosaic -- Each "Gene" has several parts. See texts or handout 25A on bottom or Purves 19.19.
B. Switching occurs at DNA and RNA levels. (Switching at DNA level is unique to immune system.)
C. How "gene" is divided -- region coding for each chain has parts for each type of constant region and several parts for variable region.
D. Region of DNA coding for variable region of H or L chain has parts called V, J and/or D.
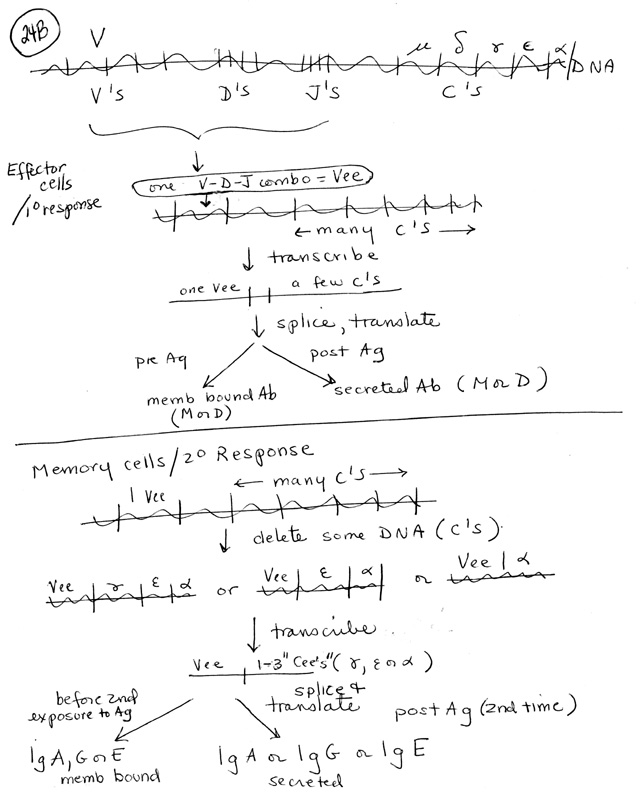
1. Pre Ag -- rearrange DNA to make one variable region per naive/virgin B. Details on handout 25B, top or Purves 19-20.
2. Post Ag -- Alt splice of RNA and/or further rearrangement of DNA (See handout 25B, bottom or Purves 19.21) --> Changes (primarily) affect constant regions of H chains. (No changes in constant regions of L chains.)
Try Problem 13-13.
IV. Details of G.O.D etc. -- Handout 25B
A. Pre Ag (Top half of handout, left fork)
1. Order of rearrangements: H rearranges first; D --> J; V --> DJ; then L rearranges (L chains are called Kappa & Lambda, have no D section)
2. Allelic exclusion (This is explained at length since it is not clear in the texts)
a. Definition: there are two copies of each gene (for H or L kappa or L lambda chains) since cell is diploid, but only one copy is expressed because only one rearranges (successfully). This is called allelic exclusion; only one allele used. Means only one L variable region and one H variable region made per cell.
b. Note this is not the usual case; if you have 2 copies of some gene for an ordinary protein (say enzyme W) then both copies (which are ready to function without rearrangement) are transcribed and used. This is why a WW has more enzyme than a Ww. (W --> active enzyme; w --> inactive enzyme)
c. Mechanism of allelic exclusion
Genes rearrange in a fixed order (first H, then L-kappa, then L-lambda etc.); if first "try" at H chain gene rearrangement is successful (rearranged allele of H codes for a functional peptide) the second H gene copy never rearranges and so is never expressed (C with promotor is too far from V). If first try fails, second copy of H chain gene rearranges; if both tries fail, cell dies. Once either copy of H gene successfully rearranges, one copy of kappa L chain gene rearranges. If all kappas fail, try lambdas, etc. So in the end, every surviving cell has one working copy of the H chain gene and one working copy of the L chain gene.
3. Making mRNA: Rearranged gene is transcribed through C-delta, than any extra sections from primary transcript are spliced out --> mRNA for IgM or IgD. (Note that IgD is made before exposure to antigen; it is not considered a secondary antibody, contrary to statement in Purves, p. 371.)
B. Post. Ag part
1. First switch (membrane bound M to secreted M) is by alt. splice (See earlier class handout or Becker fig. 19-27)
2. Class switching -- Changing C's. More DNA rearrangement at end of primary are irreversible -- remove extra C's -- see texts or Handout 25B, bottom.
3. Additional source of variation -- Somatic Mutation -- changes V, not C part.
a. As clone expands, in response to Ag, get somatic/hyper mutation in parts of DNA coding for variable regions of H and L chains. Mechanism unknown.
b. Exposure to Ag selects best variants = those that make Ab (Ab = antibody or immunoglobulin) with best binding to Ag. How selection works: Those with strongest binding are most likely to bind Ag and be stimulated to divide. Result is a kind of fine tuning of clonal expansion.
c. Somatic mutation occurs AFTER exposure to antigen; all other aspects of G.O.D. occur PRIOR to exposure to antigen.
d. Somatic mutation affects B cells (& Ig's) not T cells (& TCR's).
4. Secondary response -- Whether get G, A etc in secondary depends on Ag; stimulates dif. subset of T helpers --> dif. IL's; IL's control class switching (See texts and handout 25B bottom)
C. Summarize G.O.D -- how get so many V's?
1. H & L mix and match
2. V parts (V, D, J) mix and match
3. Joins are inexact
4. Somatic mutation (post Ag) --> changes in V, not C
Try Recitation Problems #12, problems 3 & 4.
D. TCR is similar, except no somatic mutation
E. Clonal vs. Natural Selection. Note how clonal selection and natural selection compare. In both cases, need to have many variants (diff. antibodies or dif. organisms) to be able to respond to unpredictable environmental challenges. How is this done? In both cases, make many variants and conditions select (promote propagation of) cells making the few suitable Ab (or carrying out a rare, useful function); the rest are wasted. Random generation of variants seems wasteful, but is the biological solution to preparing for change without conscious planning ahead.
V. So how do other cells get differentiated (specialized) to carry out their particular functions?
Cells of the immune system specialize (differentiate) by rearranging their DNA. Cells of the rest of the body specialize in a different way -- they keep the same DNA but they still make different proteins. How do cells with the same DNA differentiate to make different proteins? If there is time we will discuss this briefly. If you want to know more, take a look at "Lecture 26." This is provided FYI only -- the material in "lecture 26" will not be on the final.
{kind=link}
{kind=link}