C2006/F2402 -- 2005 --Outline of Lecture
#19 – Electrical Communication #3
(c) Stuart Firestein,
Columbia University,
New York, NY.
Last update
04/11/2005 02:02 PM
.
Notes by Chris Kelly
Basic Ideas
- Two fields have emerged to investigate sensory systems.
- The first is psychophysics, which studies the
relationship between the physics of a stimulus and our perception of it.
This field asks questions like: “What is the minimum light intensity that we
can perceive?” (Turns out the answer is a few photons.)
- The second is sensory physiology. The name is
narrower than the field, as this line of study also encompasses genetics and
molecular biology, &etc. The field attempts to understand the neural basis
for stimulus transduction. It asks questions like: “How do we transform
incoming photons into neural signals?”(Answer at the end of these notes.)
- Any stimulus has four central attributes:
- Modality – the nature and quality of the
stimulus … is it a visual image? A smell?
- Intensity / amplitude
- Location
- Duration
Features common to all
sensory systems
- In all sensory systems, the signaling must begin with a
primary sensory neuron, sometimes called a sensory receptor (even though we
are referring to a cell, not a protein). These receptors are
specialized for the particular sensory modality, and they are responsible for
the transformation of the physical stimulus into an electrical signal.
- The receptor cells can be either epithelial or
neuronal in origin. The hair cells in the cochlea are epithelial; whereas
the receptors in the nose are true neurons.
- Despite these developmental differences, all primary
sensory receptors are spatially polarized: they have front and back ends.
There is a detection end, and then a synaptic signaling end.
- An olfactory neuron, for example, has sensory cilia
protruding into the nasal cavity, which connect to a soma, which then
projects an axon to an olfactory glomerulus. At the glomerulus, the axon
synapses onto the next cell in line.
- A rod, found in the retina, has an outer segment
packed with photon transduction proteins. This outer segment is connected
to the cell soma, which immediately synapses on the next cell in line (a
bipolar cell). Note that a rod does not have an axon and so does not fire
action potentials –– the passive signal is enough to cause synaptic
transmission since the distance from the outer segment to the synapse is
so small. See your book for a diagram, which may be helpful.
- Auditory neurons are flask-shaped cells with
stereocilia on their sensory end. (Despite the misnomer, the “cilia” are
actually microvilli.) These stereocilia connect to a soma that makes a
synapse onto the next cell, again without an axon.
- In all of these cells, one often sees surface area
maximized at the sensory end in order to maximize the likelihood of picking
up a stimulus.
- Once these sensory neurons have captured a signal,
what is the result?
- A detected stimulus gives rise to a change in the
membrane potential of the receptor cell. This change is called the receptor
potential. These voltage changes are always graded, meaning that the
strength of the depolarization directly correlates with the strength of the
stimulus. If the depolarization is sufficiently large, an action potential
will be generated (either in that cell or in the first in line that is able,
for reasons discussed above). The farther past threshold the cell is
depolarized, the higher the frequency of action potentials that will fire as
a result.
- If the signal falls short of threshold, then no
further signaling will occur, and you (the brain) will never know about the
stimulus.
Features of stimuli
- We should revisit the four qualities of a stimulus in
more detail.
o Modality
§
The law of specific energies states that specialized receptors are
attuned to a certain kind of energy. This is generally true: photoreceptors
detect electromagnetic energy, whereas olfactory receptors are sensitive to
chemicals (smells). Remember, however, that under extreme conditions, a certain
stimulus can cause any receptor to respond. A punch in the eye results in a
flash of light, even though it is a mechanical, and not electromagnetic, energy.
§
There are four classes of receptors in humans: mechanical,
chemical, electromagnetic, and thermal.
·
Mechanical receptors are triggered by sheer physical activity. For
example, touch receptors in your skin respond to physical (somatosensory) input
because that input is sufficient to cause protein conformation change and, thus,
depolarization. (By pushing on your skin, you are literally opening channels.)
The auditory system has a complex mechanical arrangement for processing sound
stimuli. See book for details on the cochlea.
·
Chemical receptors respond to molecular ligands. The olfactory and
gustatory systems use chemical receptors.
·
Electromagnetic receptors respond to light energy. Rods and cones
are examples, since they respond to photons.
·
Thermal receptors respond to changes in temperature.
§
Receptors are attuned to a particular modality, but they are also
more finely attuned to particular qualities within that modality. For example,
all cones respond to light, but “blue” cones give maximal responses to
wavelengths of 420 nm, whereas “red” cones are tuned to wavelengths of 560 nm. A
tuning curve for a particular receptor plots the average response against a
varied parameter; for example, one could plot how much response a particular
photoreceptor gives when subjected to photons with different wavelengths. These
plots allow one to determine the stimulus parameters to which that receptor
gives the greatest response (and to which it is, hence, maximally tuned.)
§
Once the signal has passed beyond the initial receptor, how is its
modality coded? In other words, we know that by occurring in a photoreceptor
cell, an action potential must be encoding a visual stimulus –– but how do you
specify the modality once the signal has moved beyond the initial receptor cell?
·
Labeled line networks.
o
An electromagnetic receptor projects to a unique network of
neurons, so if those particular neurons are firing, it must be in response to an
electromagnetic input. Hence the labeled line –– a designated network.
o
Amplitude
§
Amplitude is typically described with dose-response curves. Along
the x-axis is the intensity of the stimulus, often on a logarithmic scale. On
the y-axis is the percentage of trials in which a subject can accurately detect
the stimulus at that intensity. The plot usually looks sigmoidal, and the
intensity for which one has a 50% accuracy rate is deemed the threshold
intensity –– the minimum intensity the stimulus must have in order to be
detected.
§
One problem with self-reported perception is that current
experiences are modulated by previous ones. For example, if you closed your eyes
and were given a one-pound weight, you would reliably report when you had
received the weight. If you were first saddled with a fifty-pound weight, it
would be hard to tell when the one pound weight had been added, even though its
magnitude had not changed.
·
The Weber-Fechner law gives the just noticeable difference (JND),
the minimum possible change in stimulus intensity that can be detected.
§
On the neural scale, how is amplitude encoded? Action potential
frequency and population frequency. The former corresponds to how often a single
neuron fires, the latter to how many neurons related to that stimulus are
firing.
o
Location
§
Determining the location of a stimulus is very important to an
organism vying to survive. It is thus a fast and robust mechanism in several of
the sensory systems. There are a few methods for encoding location.
§
(1) In many systems, each neuron has a receptive field, a
region of the environment to which it is sensitive. If a stimulus appears in a
neuron’s receptive field, that neuron will fire. One can thus know where a
particular object is based on which neurons are firing.
·
Note that the resolution of this system varies from location to
location, based on the density of receptor cells: one can reliably report
feeling two adjacent but distinct somatosensory inputs when those inputs are on
the finger, but not when they are on the back. On your back, they just feel like
a single input. The somatosensory receptors are thus denser in the fingers than
on the back.
·
Also, as networks converge, resolution goes down. If ten rods,
each with a discrete receptive field, all synapse onto the same bipolar cell
(the next neuron in the visual network), the receptive field of that bipolar
cell is equal to the sum of the fields for the rods. Resolution has thus
decreased, as the field input has been reduced to a single averaged unit,
instead of ten unique ones.
§
(2) Another mechanism for locating stimuli is based on
comparing inputs to the two main sensors (the two eyes, two ears, etc.).
·
In the visual system, we can calculate depth by comparing the
differences between the images on the right and left eyes.
·
In the auditory system, we can calculate location based on
differences in signal arrival times for each ear, as well as differences in
signal intensities. For example, if a sound comes from the left field, it will
reach the left ear before the right ear. Also, the signal will be louder at the
left ear than at the right, because of interference from the skull. These
differences can be analyzed to determine location.
§
Finally, with smell, we (3) determine location based on
gradients. We smell something and then move in some direction. If the smell has
gotten more intense, we’re going toward its source. This is a warmer/colder
location mechanism.
o
Duration
§
Some cells fire as long as the stimulus is present. Some cells
briefly fire as soon as the stimulus goes on, then stop, and then briefly fire
when the stimulus goes off. Still other cells fire continuously in response to
the stimulus, though the magnitude of the response decreases over time (within
the single stimulus presentation).
§
Let’s look closer at the last phenomenon: this is adaptation. In
some sensory systems, like smell, your perception of a stimulus weakens over
time. Why? There are biochemical reasons, but the effect can happen on the
channel, neuronal, or cortical level. There are different terms for each.
·
Desensitization is what channels do.
·
Adaptation is what cells do.
·
Habituation is what cortex does.
Adaptation
- What is the real purpose of adaptation? It can be a
long-term (i.e. seconds to hours) effect. Above, we discussed a decrease in
firing within a single stimulus trial, but adaptation can also affect one’s
processing of all trials within an environmental context. So, whereas before
we talked about neurons adapting (i.e. showing a decreased response) to the
single signal, here we are talking about neurons adapting to the environmental
context, and adjusting their responses appropriately.
- This sort of adaptation allows us to remain sensitive
over a wide range of stimulus intensities. Consider the photoreceptors, for
example. These are cells sensitive to over six orders of magnitude of stimulus
intensity. You can distinguish between objects in incredible bright rooms, and
you can do the same in very dark rooms. How can photoreceptors, despite this
incredible scope, remain highly sensitive to small changes in light? In other
words, if you plotted a response vs. intensity (dose response) curve, how can
you keep the slope extremely steep at any given point?
- If you consider your plot, instead of having a single,
monotonic increase in response based on intensity –– a line that would have
a shallow slope, given the broad domain –– you have a shifting sigmoidal
curve that allows for high changes in response for little changes in
stimulus intensity.
- What in the world am I talking about? Consider the
following graph:
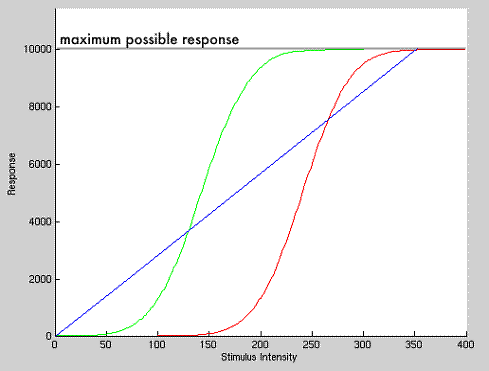
- First consider only the blue line. This line
indicates what the stimulus response curve would look like without any
adaptation –– a linear relationship between stimulus intensity and
response. But consider that the slope of this line at any point is
10,000/350=28.6. So increasing the stimulus intensity from 125 to 175
gives a response change of 28.6*50=1,430. This is okay, but we can do
better.
- Consider, now, the green line. This would be the
dose-response curve for a receptor narrowly tuned to stimuli with
intensities between 75 and 200. If we increase the stimulus from 125 to
175 on this line, we get a response change of about 4900. This is about
three and a half times greater than the change in response to the same
change in stimulus for the blue line. So the cell is much more sensitive
to changes in stimulus intensity, which is a good thing, because it means
it can easily distinguish between two stimuli presenting at similar
intensities. This is contrast, on which vision is based.
- But what about stimuli outside the range of 75 and
200? The sensitivity of the receptor, if we consider only the green line,
is extremely poor in these regions. The response is the same no matter the
stimulus! This is where adaptation comes in. Adaptation changes the
sensitivity of the neuron in question from the green line to the red line.
The neuron adapts to the new range of stimuli intensities and changes its
domain of sensitivity.
- What is a real life example? The above graph is
made-up and with arbitrary units. But imagine that it corresponds to light
intensity on the x-axis and visual response on the y-axis. If you are in a
dark room, the photoreceptors have the green response curve. You can
clearly distinguish between stimuli as long as their visual intensity is
between 75 and 200 AU (arbitrary units), since an increase of 1 AU in
stimulus intensity produces a significant change in receptor response.
When you go outside into the daylight, however, all of the stimuli are
>200 AU. Since the response is flat in this region for the green curve,
the stimuli all generate the same responses, and hence look the same. So
you cannot see anything. Through biochemical processes, though, the neuron
shifts itself to the red response curve. You are then sensitive to
differences in intensity of brighter lights and can see again. But if you
go back into the dark room, you will see nothing, of course, until the
neurons shift back to the green curve.
Transduction
- The mechanisms vary for each sensory system, but let us
consider how the visual system transforms visual signals into
neural/electrical signals.
- Within the outer segment of the photoreceptors, there is
a protein called rhodopsin. This protein was the first GPCR ever discovered,
and its G-protein (called transducin) was also the first G-protein to be
discovered.
- Rhodopsin has a molecule called retinal bound to one of
its residues. (Remember that because it is a GPCR, rhodopsin is a seven-pass
transmembrane protein.)
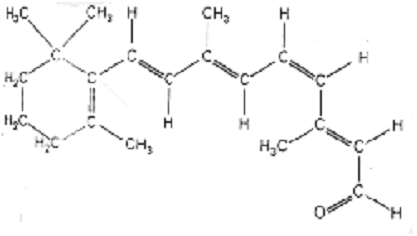
- Retinal starts in a form called 11-cis, aptly named
since the double bond at the eleventh carbon is cis. Here is a picture:
- 11-cis retinal just sits there and does nothing. When
a photon arrives at the photoreceptor, however, it gives retinal just enough
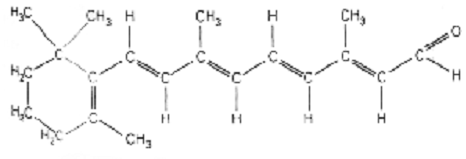
energy to isomerize its cis double bond, thereby resulting in all-trans
retinal, pictured here:
- Rhodopsin can hold 11-cis retinal, but it cannot hold
all-trans retinal without changing conformation. It has no choice, then, but
to adapt a different three-dimensional structure, which initiates the
G-protein mediated signaling cascade that results in the electrical signal
encoding vision.
- Note that only photons of certain wavelengths have the
correct energy to isomerize the double bond –– less energy is insufficient,
and greater energy is dangerous. Perhaps not surprisingly, but rather
amazingly, the photons that can isomerize 11-cis to all-trans are those
within the visual spectrum –– 400 to 700 nanometers. (If you are interested
in details on how the three different kinds of cones have different
sensitivities to different wavelengths, see your textbook.)