C2006/F2402 '05 OUTLINE FOR LECTURE #5
Last updated 02/01/05 05:04 PM(c) 2005 Dr. Deborah Mowshowitz , Columbia University, New York, NY
Handouts: 5ATransport of glucose through body -- , 5B -- Models for Active Transport , 5C--Receptor Mediated Endocytosis (5C not needed until lecture #6)
1. Kinetics and Properties of each type of Transport -- How you tell the cases apart.
A. Simple Diffusion (Case 1); Curves #1 & #2 refer to diagrams on top of handout 4A.
1. Curve #1 (uptake or concentration of substance X inside plotted vs. time) plateaus at [X]in = [X]out.
2. Curve #2 (uptake of X plotted vs concentration of X added outside) does not saturate.
3. Energy: Rxn ( X in <--> X out) is strictly reversible. (Keq = 1; standard free energy change = 0; at equil. [X]in = [X]out).
Actual free energy change and direction of transport depends on concentration of X. If [X] is higher outside, X will go in and vice versa.4. Importance. Used by steroid hormones, some small molecules, gases. Only things that are very small or nonpolar can use this mechanism to cross membranes. Materials (usually small molecules) can diffuse into capillaries by diffusing through the liquid in the spaces between the cells. (The cells surrounding capillaries do not have tight junctions, except in the brain.)
B. Carrier mediated Transport = Facilitated Diffusion using a carrier protein (Case 3)
1. Curve #1 same as above.
2. Curve #2 saturates. See Becker fig. 8-6, or Purves 5.11 (5.10 in 5th ed.; not in 6th)
3. Mechanism: Carrier acts like enzyme or permease, with Vmax, Km etc. See Becker fig. 8-8.
4. Energy as above -- substance flows down its gradient, so transport is reversible, depending on relative concentrations in and out.
5. Regulation: Activity of transport proteins can be regulated at least 3 ways. Methods a & b are common to many proteins and are only listed here for comparison (details elsewhere). Method c is unique to transmembrane proteins. (This section is about regulation of activity of pre-existing levels of protein. Synthesis and therefore protein levels are also regulated, as will be explained later.)
a. allosteric feedback inhibition/activation of carrier proteins
b. covalent modification (reversible) of the carrier proteins -- common modifications are addition and removal of phosphates; see example below (Na+/K+ pump)
c. removal/insertion of carrier into membranes.
(1). Newly made membrane proteins are inserted into the membrane of a vesicle, by a mechanism to be discussed later.
(2). Vesicle can fuse with plasma membrane; process is reversible.
(a). Fusion of the vesicle with the plasma membrane inserts transport protein into plasma membrane where it can promote transport.
(b). Budding (endocytosis) of a vesicle back into the cytoplasm removes the transport protein and stops transport.
(3). Some channels and/or carrier proteins are regulated in this way -- channel or carrier proteins can be inserted into the membrane (or removed) in response to the appropriate hormonal signals. (An example next time.)
To see how you analyze uptake, try problem 2-1. To summarize everything so far, try 2-4.
C. Channels (Case #2)
1. Curve #1 -- Same as above except
a. Very high rate of transport -- Initial slope of Curve #1 very steep.
b. Channels often conduct ions. This has consequences. Curve #1 plateaus as above with [X]in = [X]out only if X is neutral or there is no electric potential -- see point 4 below.
2. Curve #2: Shape like simple diffusion (linear, no saturation) at physiological concentrations. (Curve plateaus only at extraordinarily high concentrations, so we are assuming no saturation observed -- extremely high concentrations of X not reached.)
3. Mechanism. Lack of saturation and high rate of transport indicate that max. capacity of channel is very large and is not easily reached. This is explained by one or both of the following:
a. Binding of ion to channel protein is weak (Km >> 1), and/or
b. No major conformational change of channel protein is required for ion to pass through.
See Purves 44.5 (44.6) for comparison of ion pumps and ion channels; Becker p. 203 (209) for comparison of carrier and channel proteins. Note that channels are very specific in spite of features a & b -- each channel transports only one or a very small # of related substances. (Mechanism of specificity has been recently figured out for one channel -- see Purves 5.10 or click on link for a nice picture of a channel and an explanation of how K+ channels can be selective.)
4. Terminology. Diffusion through a channel is usually called "facilitated diffusion" because a protein is needed (to form the channel) for transport across the membrane. However, diffusion though a channel is also sometimes called "simple diffusion," because the rate of transport as a function of [X] is generally linear, as for simple diffusion, as explained in point #2 above. In other words, the kinetics of passage through a channel are linear (at physiological concentrations of X), like simple diffusion -- not hyperbolic, as in carrier mediated transport or standard enzyme catalyzed reactions. Perhaps the best term for transport through a channel is "channel mediated diffusion."
5. Gating
a. Some Channels are gated = % time any particular gate is open is controlled (but each individual gate is either open all the way or shut)
(1). Ligand gated -- opens or shuts in response to ligands (= chemicals that bind to substance under discussion). Typical substances that open ligand gated channels are hormones, neurotransmitters, etc. For a picture see Purves 5.9.
(2). Voltage gated -- opens or shuts in response to changes in voltage. Allows transmission of electrical signals as in muscle and nerve -- see Becker figs. 9-9 & 9-10.
(3). Mechanically gated -- opens or shuts in response to pressure. Important in touch, hearing and balance.
b. Some channels are open all the time (ungated); An example = K+ leak channels. These allow a little K+ to leave or "leak out" of cells, causing cells to have a slight overall negative charge. This is critical to conduction of impulses by nerve and muscle as will be explained in detail later. Why do leak channels only allow "a little" K+ to leave? See below.
7. Most channels are ion channels -- transport charged particles, not neutral molecules. This raises new energy considerations:
a. Role of charge: If X is charged, need to consider both chemical gradient & voltage (charge gradient). These can both "push" ions the same way or in opposite directions.
b. Result of charge: Keq not usually 1 here -- Curve #1 plateaus when chemical gradient and voltage are balanced (not necessarily at [X]out = [X]in). Example: K+ ions stop leaking out of the cell and you reach equilibrium for K+ when the charge difference across the cell membrane (which pushes K+ in) balances out the concentration difference across the membrane (which pushes K+ out).
See problem 2-6, A. Can you rule out transport through a channel?
D. Active Transport (Cases 4 & 5)
1. Curve #1: when it plateaus, [X]in greater than [X]out -- because movement of substance linked to some other energy releasing reaction. (This assumes we are following the reaction Xout --> X in)
2. Curve #2 saturates. Enzyme-like protein involved -- acts as transporter or pump.
3. Energy: Not readily reversible; Keq not = 1 and standard delta G not = zero. Overall reaction usually has large, negative standard delta G because in overall reaction transport of X (uphill, against the gradient) is coupled to a very downhill reaction. The downhill reaction is either
a. Splitting of ATP (in primary active transport), or
b. Running of some ion (say Y) down its gradient (in secondary active transport).
4. Secondary (Indirect) Active Transport -- How does ATP fit in? Process occurs in 2 steps:
a. Step 1. Preparatory stage: Splitting of ATP sets up a gradient of some ion (say Y), usually a cation.
b. Step 2. Secondary Active Transport Proper: Y runs down its gradient, and the energy obtained is used to drive X up its gradient. See Becker fig. 8-10.
c. Overall: Step (1) is primary active transport; step (2) is secondary and can go on (in the absence of ATP) until the Y gradient is dissipated. Note that step (1) cannot occur at all without ATP but step (2) can continue without any ATP (for a while).
5. How do you tell the two types apart? Primary is directly dependent on splitting of ATP; secondary will continue even in the absence of ATP until the gradient of Y runs down.
Try problem 2-2.
6. Some Examples & Possible mechanisms (see handout 5B, texts & animations for models). Click on links for animations.
|
Example |
Type of Active Transport |
Type of "Port" |
Pictures in Becker |
Pictures in Purves |
|
|
a. |
Primary |
Antiport |
figs. 8-11 & 8-12 (8-10 & 8-11) |
5.13 (5.12) |
|
|
b. |
Na+/Glucose co-transport |
Secondary |
Symport |
fig. 8-13 (8-12) |
5.14 (5.13) |
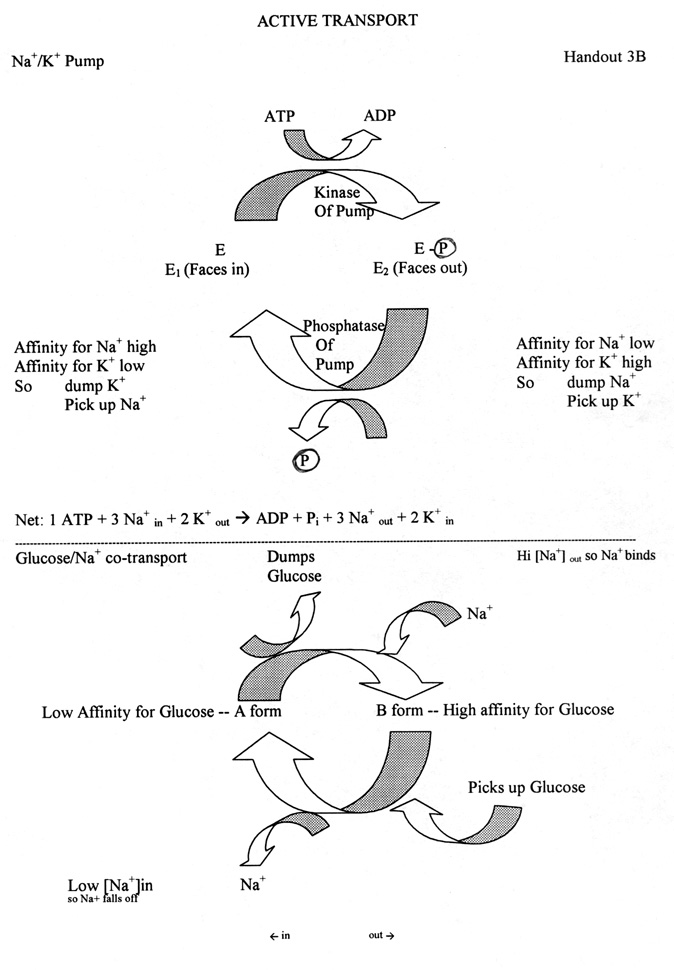
7. Important features of Na+/K+ pump (See handout)
a. Enzyme has 2 forms: one faces in (E1), one faces out (E2).
b. Forms have different affinities for K+ and Na+. (See handout)
c. Role of Phosphate: addition/removal of phosphate switches the enzyme from one form to the other.
d. Role of enzymes
(1). Use of kinases & phosphatases (for addition/removal of phosphates) is a common way of regulating enzyme activity by reversible covalent modification.
(2). Enzymatic reactions:
(a). phosphorylation -- addition of phosphate groups -- catalyzed by kinases.
Kinases catalyze: X + ATP --> X-P + ADP(b). dephosphorylation -- removal of phosphate groups -- catalyzed by phosphatases.
Phosphatases catalyze: X-P + H2O --> X + PiP (bold) = phosphate group; Pi = inorganic phosphate (in solution)
(3). Result of enzyme activity in this case:
Kinase activity (catalyzes phosphorylation) flips enzyme "out" ( E1 --> E2)
Phosphatase activity (catalyzes removal of phosphate) flips enzyme "in" (E2 --> E1)
e. Location of Enzymes: kinase and phosphatase) are part of the pump itself. Not separate proteins.
f. How one cycle goes:
Binding of Na+ on inside activates kinase. Flips enzyme out, dumps Na+, picks up K+.
Binding of K+ on outside activates phosphatase. Flips enzyme in, dumps K+, picks up Na+.
g. Stochiometry: 3 Na+ out per 2 K+ in. Some of charge differential balanced by Cl- transport. Cells are negative on inside relative to outside, but most of charge imbalance is NOT due to pump.
8. Important Features of Na+/Glucose co-transport
a. Enzyme has 2 forms with different affinities for glucose.
b. Role of Na+: Binding of Na+ switches the protein from one form to the other; alters affinity for glucose.
c. Binding of glucose probably flips protein so it faces the other way; loss of glucose does the reverse.
d. Direction of glucose transport & reversibility: Either form (with or w/o Na+) can face in or out. In normal cell, glucose always goes into the cell with Na+. Why?
9. Are pumps reversible?
a. Theoretically, all pumps (like Na+/K+ pump) are reversible -- a pump can break down ATP and use the energy to drive ions up their gradient, or (if ion gradient is large enough) ions running down their gradient can provide enough delta G to drive phosphorylation of ADP to ATP. Therefore, proteins that catalyze active transport are sometimes called "ATPases" or pumps, whether their normal function is to hydrolyze ATP or to synthesize ATP.
b. Practically speaking, inside cells, most pumps are irreversible. Most (but not all) individual "pump" proteins work only one way in cells, because the standard delta G for the "usual" direction is very negative. Therefore it takes very high concentrations of products (very high ATP or very high ion concentrations, depending on the reaction) to push the reaction in the "reverse" direction. The concentrations needed to reverse the reaction are not reached in cells, but can be achieved in test tubes (by adding ATP, setting up ion gradients, etc.). So in vitro (in test tubes), but not in vivo (in living cells), you can make the pumps run in either direction. Two examples of important pumps that are reversible (in vitro), but usually run in one direction (in vivo):
(1). In the inner membranes of mitochondria and chloroplasts, chemical or light energy is used via electron transport to set up a proton gradient, which then runs down; driving phosphorylation of ATP. So these systems almost always act to make ATP while ions run down their gradient. (Diff. proteins used in the two organelles.)
(2). The Na+/K+ pump in the plasma membrane almost always uses up ATP -- this system drives ions up their gradients at the expense of ATP.
For more examples, see Becker table 8-3.
Now try problems 2-3 & 2-5.
II. Putting all the Methods of Transport of Small Molecules Together or What Good is All This?
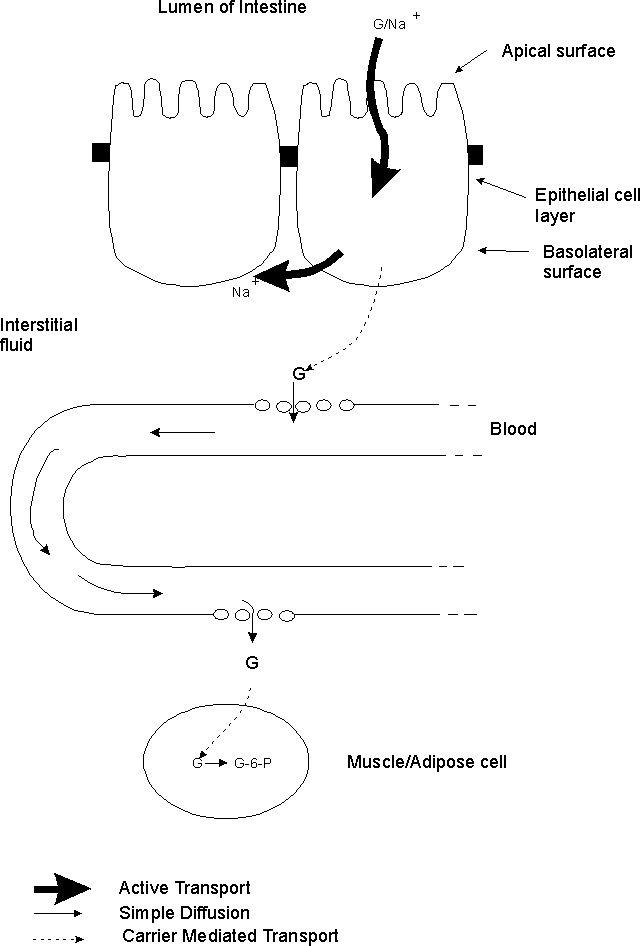
A. How glucose gets from lumen of intestine --> muscle and adipose cells. An example of how the various types of transport are used. (Handout 5A and Becker fig. 11-22) Steps in the process:
1. How glucose exits lumen. Glucose crosses apical surface of epithelial cells primarily by Na+/Glucose co-transport. (2o act. transport)
2. Role of Na+/K+ pump. Pump in basolateral (BL) surface keeps Na+ in cell low, so Na+ gradient favors entry of Na+. (1o act. transport)
3. How glucose exits epithelial cells. Glucose (except that used for metabolism of epithelial cell) exits BL surface of cell (and enters interstitial fluid) by facilitated diffusion = carrier mediated transport. (Interst. fluid = fluid in between body cells.)
4. How glucose enters and leaves capillaries -- by simple diffusion through spaces between the cells. Note: this is NOT by diffusion across a membrane.
5. How glucose enters body cells -- by facilitated diffusion (= carrier mediated transport). Carrier is only "mobilized" that is, inserted into membrane (by fusion of vesicles as explained previously) in some cell types (adipose & muscle) in presence of insulin. Carrier is permanently in cell membrane in other cell types (brain, liver). See below on GLUT transporters.
6. Role of glucose phosphorylation. Conversion of G --> G-6-phosphate traps G inside cells.
For additional examples of the uses of the various types of transport processes, see Becker fig. 8-1 & 8-2.
B. How Glucose Reaches Body Cells -- Another look at handout 5-A. -- The steps in the process are described above in the order in which they occur. Here, the focus is on the various types of transport involved.
1. Role of Active transport -- Needed to get glucose from lumen to inside of epithelial cell.
a. Primary active transport -- Na+/K+ pump keeps intracellular [Na+] low.
b. Secondary active transport -- Glucose enters epithelial cells by Na+/Glucose co-transport
2. Role of Passive Transport & Phosphorylation
a. Passive Transport -- Used to move glucose the rest of the way -- out of epithelial cells, in & out of capillaries, and into body cells.
b. Phosphorylation of glucose -- Used in the body cells to keep the free glucose level at the "end of the road" low, and ensure that the glucose gradient is "downhill" from epithelial cells to capillaries to body cells.
3. Role of Diffusion: Glucose and other small molecules (but not macromolecules) diffuse in and out of capillaries through the liquid filled spaces between the cells, not by diffusing across the cell membrane. Note that proteins are too big to enter or leave capillaries this way.
4. Role of GLUT transporters (another protein/gene family)
a. GLUT proteins are responsible for carrier mediated transport of glucose. All passive glucose transport across membranes depends on a family of proteins called GLUT 1, GLUT 2, etc. This family of genes and transport proteins is responsible for all carrier mediated transport of glucose.
b. Different family members (genes and proteins) are expressed in different cell types. GLUT 1 protein is found in plasma membrane of RBC & most other cells, GLUT 2 protein on BL surface of intestinal epithelial cells, GLUT 4 protein in muscle and adipose, etc. (Note all genes for all proteins are present in all these cell types -- DNA is the same!)
c. All the genes and corresponding proteins are similar, but have significant structural and functional differences. This is another example of a gene/protein family. All the proteins have a similar overall structure -- 12 transmembrane segments, COOH and amino ends on intracellular side of membrane, etc.
d. Position & Action of GLUT 4 is insulin dependent. GLUT 4 is the only insulin dependent member of the family. Insulin triggers insertion of GLUT 4 protein into the plasma membrane, by triggering vesicle fusion, as explained above. All the other proteins are located constitutively in their respective membranes.
e. Direction of transport. Note that one member of this family (GLUT 2) is responsible for ferrying glucose OUT of epithelial cells; different members are responsible for helping glucose ENTER most other cells. All family members bind glucose on one side of the membrane, change conformation and release glucose on the other side of the membrane. Which way the glucose goes (overall) depends on the relative concentrations of glucose on the two sides of the respective membrane, not on which GLUT protein is used.
Try problem 2-9.
Next time: How do large molecules cross membranes? Then how do newly made proteins get to the right place?
{kind=link}
{kind=link}
{kind=link}