C2006/F2402 '06 -- Outline Of Lecture #3 -- Last update 01/23/2006 01:52 PM
© 2006 Deborah Mowshowitz, Department of Biological Sciences, Columbia University, New York NY
Reminder: SURF symposium is next Wednesday, Feb. 1. See link for details and how to earn a little extra credit.
Handouts:
3A
-- Epithelial cells (Examples of Cell-Cell connections) & RBC Membrane
3B -- Cell-Cell Junctions, ECM, & types of Membrane Proteins
I. Membrane Structure --
Fluid Mosaic Model
A. Fluid Part = Lipid bilayer
1. Amphipathic nature of lipids (See Purves 5.2) -- there are multiple different "two headed" lipids -- each type has a different structure, but each has a hydrophobic end and a hydrophilic end.
2. Lateral diffusion vs. flip-flop. See Becker 7-10 & 7-11.
lateral diffusion = movement within plane of membrane -- fast (secs). Animation of lateral diffusion.
flip-flop = movement from one side of bilayer to the other -- slow (hrs) w/o enzymes. Need enzymes (flippases) to speed flip flop.
3. Two sides of bilayer (leaflet) can have different lipids.
B. Mosaic Part = Protein. Types of Membrane Proteins -- what do you get if you take a membrane apart?
1. Peripheral membrane proteins vs. integral membrane proteins
|
Type of Membrane Protein |
Alt. terminology |
Protein Removed From Membrane By |
Location/Attachment of Protein |
|
Peripheral |
Extrinsic |
salt, pH changes |
On one 1 side of bilayer; non covalently attached to lipid |
|
Integral |
Intrinsic |
disrupting lipid bilayer |
Goes through bilayer* or Covalently attached to lipid on one side (Lipid-anchored)** |
* A small number of integral proteins do not go all the way through the membrane; they will be largely ignored in this course. For examples see Becker fig. 7-19 (first protein on left) or Purves 5.1 (last protein on right).
**Note that lipid-anchored proteins can be considered a type of integral protein or a separate category. See Becker fig. 7-19.
2. Transmembrane proteins (See Purves 5.4 and/or Becker fig. 7-19 & 7-21)
a. Single pass vs multipass
b. Domains -- intracellular, extracellular, transmembrane
c. Location of carbohydrates -- all in extracellular domain
d. Anchorage -- Some proteins are anchored to cytoskeleton; some float in lipid bilayer.
e. Functions -- Bridge the membrane -- facilitate transport of materials & signals across membrane; physically connect cytoskeleton (inside of cell) to materials on outside of cell (ECM = extracellular matrix) or to next cell.
II. The Red Blood Cell (RBC) Membrane -- The best studied example of a Membrane.
A. Why RBC's
1. Easy to get
2. No internal membranes -- all organelles lost during maturation of RBC -- see Becker fig. 7-20 (a). Only membrane = plasma membrane.
3. Can make ghosts = resealed plasma membranes. Can be resealed (or broken and reformed into vesicles) in either orientation -- "right" or "wrong" side out.
B. RBC membrane proteins -- Structure & Function. See Purves fig.4-23 (6th ed.) or Becker fig. 7-20 (b) & 15-22 (22-22). (Handout 3A -- bottom)
1. Peripheral proteins -- spectrin, ankyrin, (band 4.1), actin. Comprise peripheral cytoskeleton, which supports membrane. All cells thought to have a similar structure under the plasma membrane.
2. Intrinsic proteins
a. Examples
(1). Multipass (band 3/anion exchanger) -- Catalyzes reversible exchange of the anions HCO3- (bicarb) and Cl- between RBC and plasma. Exchange allows max. transport of CO2 in blood (as bicarb in solution). See Purves 48.14 ( 48.17) or Becker 8-3.
(a). Basic point: Bicarb is much more soluble in plasma than CO2, so lots of bicarb (but not much CO2) can be carried in the blood. So need to covert CO2 to bicarb when want to carry CO2 in blood; need to do reverse to eliminate the CO2 (in lungs).
(b). Conversion of CO2 to bicarb (& vice versa) can only occur inside RBC, where carbonic anhydrase is. Carbonic anhydrase catalyzes CO2 + H2O ↔ HCO3- + H+
(c). Gases can pass through membranes by diffusion -- CO2 can exit or enter RBC as needed. However bicarb cannot pass through membranes. You need the anion exchanger to get bicarb in and out of RBC.
(d). Where CO2 is high, as in tissues, CO2 diffuses into RBC and is converted to bicarb inside the RBC. Then bicarb leaves RBC in exchange for chloride using the anion exchanger.
(e). In lungs, the process is reversed -- bicarb reenters the RBC in exchange for chloride using the anion exchanger. The bicarb is converted back to CO2 inside the RBC and then the CO2 diffuses out of the cells and is exhaled.
(2). Single pass (glycophorin) -- function of protein not known.
(a). Large amount of (-) charged carbohydrate -- sialic acid -- may cause RBC to repel each other and prevent clumping of RBC.
(b). Loss of terminal sugars may occur with age and trigger destruction of "old" RBC.
(c). Glycophorins make up a gene family; variations in glycophorin A are responsible for MN blood type differences. Variations in glycophorin C lead to resistance to malaria.
Try problems 1-2 & 1-3. To review membrane structure, try 1-15 to 1-18.
III. Extracellular Matrix (ECM)
Note: Becker Chap. 17 (11) goes well beyond what will be covered in this section. References to pictures and diagrams are included FYI.
A. Major Structural Proteins -- collagen -- nice picture in Becker fig. 17-2 (11-2) and elastin (diagram in Becker fig. 17-4).B. Adhesive Proteins -- fibronectins, laminins, etc. Have multiple binding domains. Connect other materials in ECM with each other and/or membrane proteins. For pictures see Becker figs. 17-6 & 17-7.
C. Proteoglycans -- special type of glycoprotein consisting of lots of carbohydrate attached to a protein core. Provides a gel-like matrix for ECM. See Becker fig. 17-5 (11-5 & 11-6). Table below is for reference purposes only so you can follow the terminology. See http://www.indstate.edu/thcme/mwking/glycans.html for a nice web site with a summary of structure, function, and medical significance of proteoglycans and GAGs.
|
Proteoglycan (mucoprotein) |
Glycoprotein |
|
|
General description |
Lots of carbohydrate attached to a protein Core;* Can be 95% carbohydrate |
A protein with some Carbohydrate Attached |
|
Are sugar chains branched? |
No |
Yes |
|
Length of Sugar Chain |
Long |
Short |
|
Are sugars repeating? |
Yes (repeating disaccharide) |
No |
|
Type of Carbohydrate |
Mucopolysaccharide or GAG (glycosoaminoglycan) |
Oligosaccharide |
|
Example |
See Becker fig. 17-5 (11-5). |
Band 3 protein or glycophorin |
|
Location |
Extracellular matrix (form gel) |
Integral membrane protein (carbohydrates on extracellular domain) |
* Multiple proteoglycans can be attached to a core carbohydrate chain (GAG or mucopolysaccharide) to form a giant aggregate as shown in Purves fig. 4-26 (4-28) or Becker fig. 17-5 (11-6). (An individual proteoglycan is shown in Becker fig. 17-5 (11-5).)
** Widely used as an anticoagulant. Inhibits factor required for blood clotting. (Physiological role, meaning real job in body, may not be as anticoagulant. Is used to remove fat particles from blood after a fatty meal.)
D. Basal Lamina (see Becker 17-8 (11-9) & epithelial cells below.)
1. Structure -- Solid layer found in parts of ECM. Main components are networks of laminin & collagen. (For structure of laminin, see Becker fig. 17-9 (11-10))
2. Location -- surrounds some cells (skeletal muscle, fat) and underlies some epithelial layers (on basal side).
3. Terminology -- Also called basement membrane especially in older literature. Has no lipid & is not a real membrane.
4. Function -- physical barrier, support and/or filter.
5. How Connected to cells -- see integrins and hemidesmosomes below.
E. Connection to cytoskeleton -- ECM often connected to transmembrane proteins called integrins. Integrins link ECM and cytoskeleton. More details below.
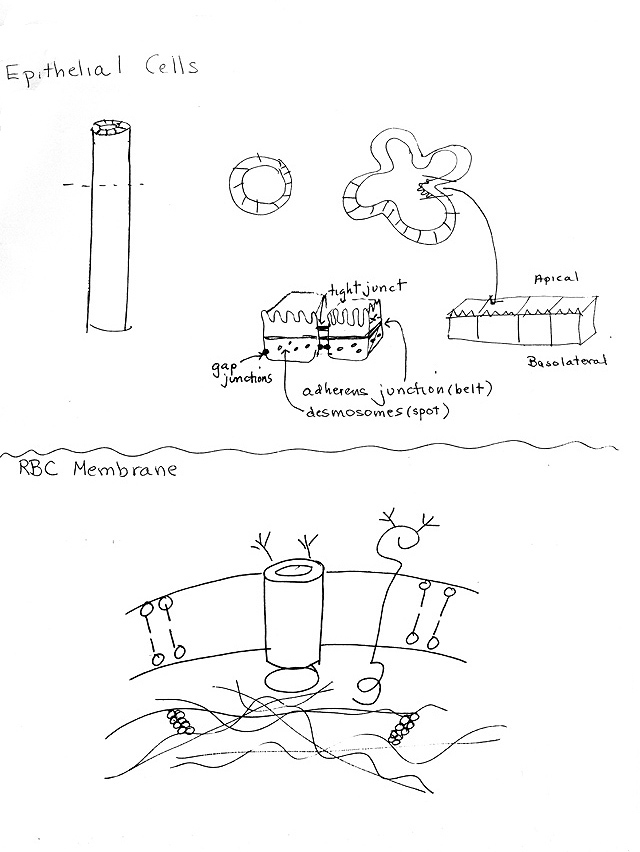
IV. Cell-Cell (& Cell-ECM) Connective Structures -- For Pictures see Purves 5.6, & Becker Ch. 17 (11) --exact figures listed below. For comparisons, see table below and/or Becker table 17-3 (11-3).
A. Cell-Cell Junctions -- for overall view, see Becker fig 17-17 (11-18). For the molecular details (FYI only) see fig. 17-12 (11-13) .
1. Gap junctions -- Becker fig. 17-21 (11-23); Purves 5.6 & 15.16 (15.18).
2. FYI: Plasmodesmata -- Purves 15.17 (15.19) or Becker 17-25 (11-28). The plant's equivalent of a gap junction.3. Tight Junctions -- Becker fig. 17-19 (11-20)
4. Adhesive (anchoring) junctions -- Spot (with IF) = desmosomes vs belt (with MF) = adherens junctions. Becker fig. 17-18 (11-19) & 17-13 (11-14) for molecular details (for your interest only). For a nice EM picture see http://trc.ucdavis.edu/mjguinan/apc100/modules/Integument/_index.html (click on generic desmosome).
|
Summary Table of Animal Cell-Cell Junctions: |
||
|
Name of Junction |
Important Structural Features |
Function |
|
Gap Junction |
Connexons; small gap between cells (2-4 nm) |
Passage of small molecules and ions (signaling & nutrition) |
|
Tight Junction |
Fusion of ridges of membrane -- no gap at ridge. |
Water Tight Seal between cells; divide membrane into regions |
|
Adhesive or Anchoring |
Intracellular Plaques with filaments. Classified
by: |
Strength |
B. Cell - ECM junctions. Connect cytoskeleton to ECM or solid support. Resemble half of an adhesive junction (adherens junction or desmosome).(Becker Fig. 17-11 (11-12). Diagrams in 5th ed. are not quite right but pictures are good. Diagrams look ok in 6th.) For a better diagram see http://celljunctions.med.nyu.edu/images/figure1.gif. For a nice EM picture see http://trc.ucdavis.edu/mjguinan/apc100/modules/Integument/skin/hemidesmosome/hemidesmosome2.html.
1. Similarities to cell -cell adhesive junctions
a. Plaque (thickening) forms on inside of cell near junction
b. Plaque connects transmembrane protein to IF or MF on inside of cell.
2. Differences from cell-cell adhesive junctions
a. Transmembrane protein is an integrin, not a cadherin (See Becker fig. 17-10 (11-11) for a diagram of structure of integrin.)
b. Transmembrane protein connects to ECM. (Not to extracellular domain of protein from another cell. )
3. Two Types.
a. Hemidesmosomes = "half desmosome"; connect to IF on inside of cell. Connect to basal lamina on outside.
b. Focal Adhesions = "half adherens junction" -- connect to MF on inside of cell and ECM (can be on a solid support) on outside.
At this point, it is a good idea to make a chart for yourself that classifies (or compares and contrasts) all the types of junctions, whether cell-cell or cell-ECM. Also try problems 1-12 & 1-14.
V. What does a real cell look like? Where are the junctions, etc.? -- Becker Fig. 17-17 (11-18) or handout 3A, top.
A. Cells & Tissues
a. Specialization. All cells in multicellular organism are specialized; there is no "typical cell."
b. Types. About 200 different cell types per human.
c. Tissue = Group of cells with similar structure & function that work as a unit.
d. 4 Major cell/tissue types -- muscle, nerve, connective, epithelial
e. Terminology Note: "tissue" is also used in a nonspecific way to mean a group of cells derived from an organ or system as in "kidney tissue." A kidney is an organ made up of many different tissue types.
B. The Four major Tissue Types (See Purves 41.2 (40.2))
a. Muscle -- three kinds (skeletal, smooth & cardiac muscle); all specialized for contraction.
b. Nervous -- individual cell is called a neuron. Specialized for conduction of messages.
c. Connective -- cells dispersed in a matrix. Extracellular matrix can be solid (as in bone), liquid (as in blood) or semi-solid (gel like) as in cartilage, adipose. (Note fat in adipose is stored inside the adipose cells in vesicles, not between cells in the matrix.) See Purves 40.4 in 6th ed. Surround other tissues and provide support, protection and transport of materials to and from tissues (in blood).
d. Epithelial -- see handout 3A -- example of cells with many types of junctions
1. Cells tightly joined
2. Make up linings of external and internal surfaces
3. Usually sheets. Can have one or more layers
4. Often rest on noncellular support material = basal lamina = part of ECM secreted by cells
5. Can form glands = epithelial tissues modified for secretion. (More details on formation, structure & function of glands later in course.)
6. Usual functions: selective absorption (selective transport across sheet), protection, secretion (from glands)
7. An example: epithelial layer surrounding the gut. See handout 3A and Becker fig. 17-17 (11-18) or Purves 41. 3 (40.3) .
This leads to the next topic: How does the intestinal epithelium (& how do various types of junctions) function in transport?
Now try problem 1-13. By now you should be able to do all the problems in problem set #1.
{kind=link}
{kind=link}