C2006/F2402 '06 OUTLINE OF LECTURE #7
(c) 2006
Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last update 02/06/2006 02:32 PMHandouts: 6B -- Signal Hypothesis -- Co-translational Import ; 7A = How proteins insert into ER membrane 7B = Golgi & Lysosomes; not on web.
I. ER -- How does Co-translational Import Work?
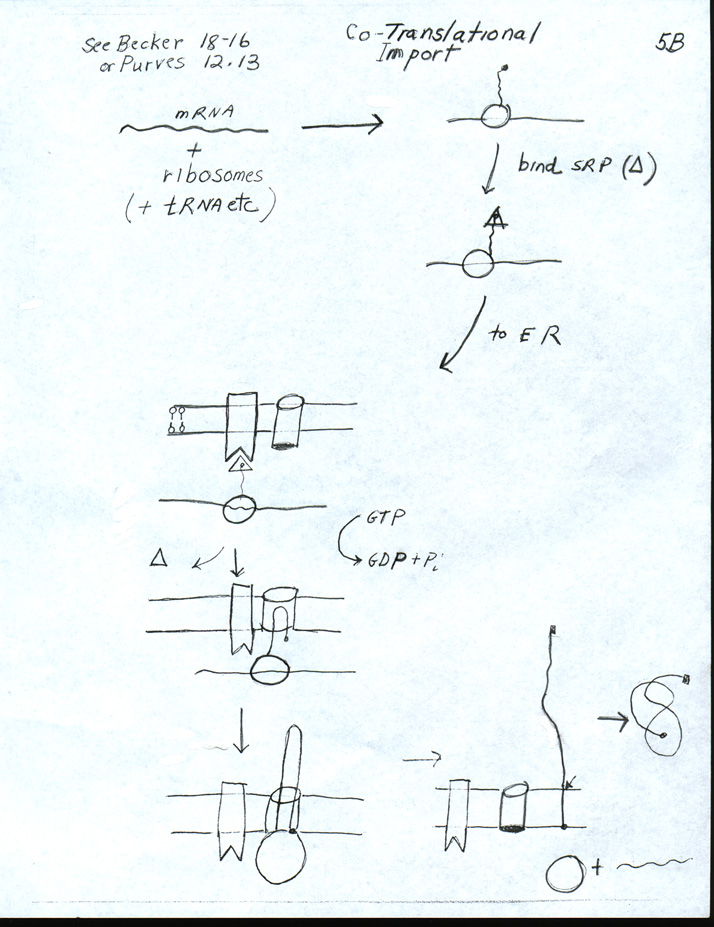
(This is a repeat of section V of notes of #6.)A. Signal hypothesis -- How ribosomes get to the ER & Protein enters ER -- Handout 6B. Steps below refer to handout. See Becker fig. 22-16 (20-16) or Purves 12.15
b. SRP = signal recognition particle1. What is the Signal Hypothesis? Ribosome unattached to ER starts making protein. (step 1) If nascent (growing, incomplete) peptide has a "signal peptide," then ribosome plus growing chain will attach to ER membrane, and growing chain will enter ER as it grows.
2. How does ribosome get to the ER?
a. Signal peptide (SP) = section of growing peptide (usually on amino end) binds SRP. (step 2)
3. How does growing chain enter the ER?
a. Ribosome/translocon complex formation occurs. (step 4)
- SRP is released, recycles -- GTP split
- Ribosome binds to pore/translocon
- Translocon opens & peptide enters (as loop).
- Ribosome resumes translation.
b. Translation (and movement through translocon) continues (step 5) Handout shows how a soluble protein ends up inside the lumen of the ER. How a transmembrane protein -- single pass or multipass -- ends up inserted in the ER membrane will be explained next time.
c. Translation (and translocation) are completed, translocon closes. (step 6)
e. Signal Peptidase (usually) cuts off signal peptide at arrow. (step 7)
- Protein is released into lumen of ER
- Ribosome and mRNA are released
Now try problem 3-4, especially part D. (If some of the parts are not obvious, wait until later.)
B. How do proteins cross the ER membrane? (See handout 7A, bottom of 6B, and/or fig. 22-17 (20-17) of Becker)
1. How proteins enter/pass through the membrane -- important points
a. ER membrane contains gated pore (also called channel or translocon) for the growing chain.
Note on terminology: The term "translocon" is used in (at least) 2 different ways. Sometimes used to mean only the channel itself, and sometimes it is used to mean the whole complex of proteins required for translocation of proteins across the membrane -- the channel, SRP receptor, etc. It should be clear from context which usage is meant at any one time.
b. SP probably forms loop not arrow. Loop enters channel (translocon) in membrane. SP loop is probably what opens (gates) the channel on the cytoplasmic side.
c. Growing protein chain passes through channel as chain is synthesized.
d. Where will protein end up? Protein can go all the way through the membrane and end up as a soluble protein in the lumen (as in example above) or protein can go part way through and end up as a transmembrane protein.
e. How do transmembrane proteins get anchored in the membrane? A hydrophobic sequence may trigger opening of the pore sideways, so protein slides out of pore, laterally, into lipid bilayer. These hydrophobic sequences are called 'stop-transfer' sequences and/or 'anchor' sequences.
2. Types of Proteins that can result (see handout 7A)
a. Soluble protein in lumen. Happens if protein passes all the way through the membrane and SP (on amino end) is removed, as above.
b. Integral membrane protein anchored in membrane by SP with no cytoplasmic domain. This happens if SP is on the amino end and is not removed.
c. Single Pass transmembrane protein -- get one of 2 possibilities:
(1). Amino end on outside of cell (= on lumen side).
One way this could happen: If SP is on amino end, and SP removed, and there is a hydrophobic sequence (acting as a stop-transfer sequence) in the middle of the peptide.(2). Carboxyl end on outside.
One way this could happen: If SP is in the middle, not on amino end. SP in this case is not removed -- it becomes the transmembrane domain of the protein. (SP doubles as stop-transfer sequence.)d. Multipass transmembrane protein. (Requires one SP and several hydrophobic (start/stop) sequences.
(1). Hydrophobic sequences can stop the process (of moving through pore) and anchor protein in membrane, as explained above.
(2). Hydrophobic sequences in the middle of the peptide can restart looping → multipass protein. These are usually called "start-transfer" sequences.
(3). 'Start-transfer' and 'stop-transfer' sequences are probably equivalent. Role depends on where in protein they occur.
(4). Every SP is also a "start transfer sequence" but not every start/stop sequence is necessarily an SP. To act as an SP, a sequence must bind to the SRP. If a hydrophobic sequence does not bind the SRP, it will not direct ribosomes to the ER.
e. Lipid Anchored Proteins: Proteins to be anchored to lipids on the outside of the plasma membrane are inserted into the ER. After the protein reaches the plasma membrane, the extracellular domain is detached from the rest of the protein and attached to lipid. (Proteins to be anchored to the plasma membrane on the inside are made on cytoplasmic ribosomes.) See Becker if you are curious about the details.
Now finish problem 3-1 & try 3-2, A -C. To test yourself on structure/insertion of integral membrane proteins, try problem 3-3.
II. What Else Happens in/on the ER?
A. What happens inside ER
1. Terminal SP usually removed by signal peptidase (enzyme) inside ER.
2. Folding of protein -- requires chaperones.
a. Chaperones (also called chaperonins) -- proteins needed to assist in protein folding. Chaperones are used every time a protein remains unfolded or becomes unfolded to cross a membrane (or refolds on the other side). Two or more types of chaperones are involved in protein folding-- different ones are found in different parts of the cell.
b. Chaperones are of two major types (families) -- HSP 60 or HSP 70 depending on molecular weight (60 K vs 70 K) and mode of action. (See an advanced text if you are curious about the mechanisms.)
c. The major chaperone inside the ER is a member of the HSP 70 family, also called "BiP"
d. HSP = heat shock proteins = chaperones; these proteins were originally discovered because they are made in large amounts after exposure to high temperatures.
3. Enzymatic Modifications. The appropriate enzymes catalyze the following:
. In eukaryotes, all S-S bonds are formed in proteins inside the ER. Proteins made in the cytoplasm do not have S-S bonds. Cytoplasmic proteins do contain cysteines and have free SH groups.a. Making of S-S bonds
b. Start of N-glycosylation (oligosaccacharides are added to the N of the amide of asparagine side chains -- called N glycosylation.) See Becker fig. 12-7 if you are curious about the biochemical details.
4. Some proteins stay in ER (in lumen or membrane); most move on to Golgi.
5. What happens to proteins in ER that do not fold properly? See Becker p. 755-757 (733-734).
a. Transport to cytosol -- Unfolded proteins are transported back to the cytosol (through the translocon -- mechanism unknown).
b. Ubiquitin addition -- in cytosol, proteins to be degraded are marked for destruction by addition of a small protein called ubiquitin. (See Becker or advanced texts if you are interested in the enzymatic details.)
c. Degraded by Proteasome -- proteasome = a large protein complex in cytosol that degrades ubiquitylated proteins to fragments, at expense of ATP. Major site of degradation of cellular proteins.
d. What goes to the proteasome? Proteins that are misfolded, damaged, or have served their function.
A major proportion of all proteins made in cell do not fold properly and are degraded.
Destruction of many proteins is regulated -- level of protein activity can be controlled by protein degradation as well as by rate of synthesis, feed back of activity, modification, etc. More details and/or examples to follow.
B. What happens on outside of ER (besides protein synthesis)
1. Lipid synthesis --
(exchange) proteins.a. Insertion: Lipids made and inserted on cytoplasmic side of membrane by enzymes attached to/in membrane.
b. Flipping: Enzymes ('flippases') required to move amphipathic lipids from one leaflet (P side) of membrane to other leaflet (E side).
c. Transport: Lipids can reach parts of cell not connected to ER through vesicles and/or transport
2. Detoxifications and other reactions catalyzed by proteins on cyto side of ER. See text for details if interested.
To review the structure and function of the ER, try problem 3-4.
III. Golgi Complex -- Structure & Function A. How things get there -- from ER in coated vesicles (coat made of protein called coatomer or COP instead of clathrin). For animation of how materials pass from ER to and through Golgi click here. B. Structure & Overall Traffic Flow - See Purves, fig 4.12 or Becker fig. 12-4 & 12-8, and handout 7B.1. Two sides of stack
a. cis/forming face (side closest to nucleus & ER)
b. trans/maturing face (away from nucleus)
2. Three basic parts or compartments in a stack
a. CGN (cis-Golgi network) or cis Golgi
b. medial cisternae (sacs) -- part in between 'cis' and 'trans' Golgi
c. TGN (trans-Golgi network) or trans Golgi
2. Different marker enzymes/functions found in different parts. (See Becker figs 12-5 & 12-6) Enzymes unique to any one cell organelle or compartment are called 'marker enzymes' = their presence is a 'marker' for the presence of that compartment or organelle.
3. Sacs in stack connected by vesicle traffic -- not completely clear which way transport vesicles go or what they carry. It is clear that newly made protein and lipid passes through the Golgi from the cis face to the trans face, and that vesicles from one stack can fuse with another in vitro. Models for transport through the Golgi are discussed in more detail later. (If no time today, we'll discuss them next time.)
C. Function -- what reactions take place inside Golgi?
-- oligosacch. that was added to glycoproteins in ER is modified. Oligosacch. is attached to "N" of amide side chains of asparagines (asn's).1. Finish N glycosylation
2. Do O glycosylation of glycoproteins. Sugars are added to "O" of the hydroxyl of the side chain of ser & thr.
3. Assemble sugars of proteoglycans (linear chains of repeating sequence = GAGs)
4. Concentrate, sort proteins. This occurs at trans face (TGN). Different areas of Golgi have receptors that trap proteins going to different destinations.
To review how proteins are directed to the right place and modified in the ER and Golgi, try problem 3-2.
IV.
Transport Through the Golgi -- Are cisternae stationary or do they progress?
A. Background
1. Transport vesicles can bud off one sac (cisterna) of the Golgi and fuse with another. However it is not completely clear which way the transport vesicles go or what they carry.
a. Direction of vesicle traffic: is it cis to trans = forward = anterograde, or vice versa = retrograde? (Older models assumed it was forward, but current evidence indicates it is both -- some vesicles go in one direction and some the other, as shown in Becker fig. 12-8.)
b. What do the vesicles carry: Is it newly made proteins from the ER and/or enzymes to modify the new proteins?
2. Net Direction of Transport. It is clear that newly made protein and lipid pass through the Golgi from the cis face to the trans face.
3. Vesicles from one stack can fuse with cisternae of another stack in vitro. (See problem 3-10.)
B. Models
1. "Vesicle Transport Model" or Stationary Cisternae Model
a. Transport vesicles move primarily forward (towards trans face).
b. Vesicles carry newly made proteins (from the ER) to next part of Golgi for additional modifications.
c. Sacs of Golgi (with their characteristic enzymes) stay put, and newly made proteins pass from sac to sac by means of vesicles.
d. Enzyme composition of each sac stays the same. Each part stays in the same place and holds on to its characteristic enzymes. It is the substrates of the enzymes (the newly made proteins) that pass through, carried by the vesicles from sac to sac.
2. "Cisternal Maturation Model" = a modified Progression Model
a. Transport vesicles move primarily retrograde (towards cis face).
b. Vesicles carry enzymes to modify and sort proteins. They do not carry the newly made proteins from the ER.
c. Sacs of Golgi move, carrying newly made proteins & lipids inside. New sacs are constantly formed at the cis face from material transported from the ER. Old sacs are lost from the trans face as they age.
d. Enzyme composition of each sac changes with time. The enzyme composition of each individual sac is constantly changing as it ages and passes from cis to trans face. However the characteristic enzymes found in the sacs at each position of the Golgi (cis, medial & trans) remain the same, because the enzymes are "passed back." The vesicles retrieve the enzymes of "older" sacs (closer to the trans face) and carry them back to newer sacs (closer to the cis face).
3. Connection Model -- another possibility is that the Golgi sacs are actually connected (although they seem to be separate), and that proteins move back and forth from one sac to the next, although net bulk flow of newly made proteins is from cis to trans. Some recent data using genetically modified protein tagged with GFP has been interpreted in support of this model.
4. What really happens? The models above are not mutually exclusive, so a hybrid model is possible. Only new experiments generating new data will settle the question of how material actually moves. Not all materials may move through the Golgi in the same way. For a review of the models and the data as of 1998 (the 100th anniversary of the discovery of the Golgi) see Coming to Grips with the Golgi.
To review traffic through the Golgi, try problem 3-10.
Next Time: Sorting in the TGN: How do normal hydrolases get to lysosomes? What happens if (a) one hydrolase is not made or (b) all the hydrolases lack M6P? How do enzymes get to peroxisomes?
{kind=link}
{kind=link}