C2006/F2402 '08 OUTLINE OF LECTURE #11
(c) 2008 Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last update 03/04/2008 02:49 PM .
Handouts: Handouts are 11A (Big Bang Model of Signaling), 11B (Alternative Splicing)
Links to diagrams and animations on signaling
are at
http://www.columbia.edu/cu/biology/courses/c2006/links04/signallinks.html.
These links are from previous years but should still be useful. If you find
other good ones, let me know.
I. Regulation of Eukaryotic Gene Transcription, cont.
This replaces section III-C-2, f & g, of lecture 10. I have not repeated anything that was included for reference only, but everything else is included here.
Structure & Function of Regulatory (= Tissue Specific) TF's, cont.
→ homodimer1. Co-ordinate control. A group of genes can all be turned on or off at once in response to the same signal (heat shock, hormone, etc.).
a. Prokaryotes vs. Eukaryotes: Both prok. and euk. exhibit co-ordinate control, but mechanism is different. (See table in lecture 10.)
b. Location of coordinately controlled genes
(1). In prokaryotes, coordinately controlled genes are located together in operons.
(2). In eukaryotes, coordinately controlled genes do not need to be near each other -- they just have to have the same control elements. See Sadava 14.16 (14.14).
c. Control elements:
(1). Coordinately controlled genes have the same control elements. All genes turned on in the same cell type and/or under the same conditions have the same control elements -- therefore these genes all respond to the same regulatory TF's. Result is multiple mRNA's, all made in response to the same signal (s).
(2). Most genes have multiple control elements. Transcription of any particular gene depends on the combinations of
(a). TF's (trans-acting factors) available in that cell type
(b). The control elements (cis acting regulatory sequences) in the DNA for that gene.
d. Differences in TF's. Different cell types make different regulatory TF's. Therefore different genes are turned on/off. See Becker fig. 23-24.
e. Eukaryotes vs Prokaryotes. For a comparison of regulation of transcription in prokaryotes vs multicellular eukaryotes -- see table in lecture 10.
2. Modular Structure of Regulatory TF's
a. Each TF has multiple domains.
(1). Each TF has a transcription regulation domain (also called 'trans acting domain' or in many cases 'transcription activating domain'). This domain determines the effects of DNA binding by a given TF (activation vs repression of transcription).
(2). Each TF has a DNA-binding domain -- specific for particular DNA sequence(s) and/or gene(s)
(3). TF's may have additional domains. Some examples:
(a). TF's that are hormone receptors also have a hormone-binding domain.
(b). Dimerization domains. Many TF's must dimerize to work. (Monomer is inactive.)
Some TF's form dimers with other molecules of the same protein
Some TF's form dimers with a different protein → heterodimer
(4). Order of 3 basic domains -- in hormone receptor TF's the order of domains is always the same, reflecting common ancestry (homology):
activation domain -- DNA-binding domain -- hormone binding domain
b. Equivalent Modules (domains) can be switched -- Recombinant DNA methods can be used to make hybrid TF's. This has allowed us to figure out what domain does what, and has many uses in research. Some examples are in later problem sets (for example, 6-19).
c. How do domains of regulatory TF's act?
(1) DNA binding domains -- bind to control elements (distal or proximal) in genome
(2) Transcriptional regulation domains -- bind to other proteins. "Other proteins" can be basal TF's, other regulatory TF's, or co-activator or co-repressor proteins
d. Types of DNA-binding domains. See lecture 10 (this topic is for reference only)
To review transcription, try problems 4R-5 and 4R-6A.
II. Domains & Motifs in General -- This section was included in the previous lecture for reference only. See Section III of lecture 10.
III. Overall Regulation of Eukaryotic Gene Expression -- What has to be done to make more or less of a protein? A different protein? What steps can be regulated? (This = topic IV of previous lecture.)
A. In prokaryote (for comparison) -- process relatively simple.
1. Most regulation at transcription.
2. Translation in same compartment as transcription; translation follows automatically.
3. Most mRNA has short half-life.
B. In eukaryote -- Gene expression has many more steps & complications than in prokaryotes -- more additional points of regulation -- not just at transcription. See Becker fig. 23-11 (21-11) or Sadava 14.12 (14.11).
C. How can amount of protein be controlled? If cell makes more or less protein, which step(s) are regulated?
1. Transcription is main point of control, but
a. Need to unfold chromatin first to make transcript. (See above.)
b. Once transcript is made, it is not automatically translated. (See below.)
2. Transcript must be processed (capped, spliced, polyadenylated, etc.) -- any of these steps can be regulated, and there is more than one way to process most primary transcripts.
3. Transcription & translation occur in separate compartments.
a. mRNA must be transported to cytoplasm.
b. Translation can be regulated (independently of transcription) -- can control usage and/or fate of mRNA, not just supply of mRNA. For any particular mRNA, can regulate 1 or both of following:
(1). Rate of initiation -- can control how often ribosomes attach and start translation.
(2). Rate of degradation -- can control half life of mRNA. Some mRNA's are long lived and some have a very short half life.
D. If cells make different proteins, how is that controlled? If two eukaryotic cells (from a multicellular organism) make different proteins, what is (usually) different between them?
1. Is DNA different? (No, except in cells of immune system.)
2. Is state of chromatin usually different? (Ans: yes) How is this tested? Method & result described previously. See figure 23-17 (21-17) in Becker.
3. Is mRNA usually different? (Ans: yes). This means you can get tissue specific sequences from a cDNA library. (cDNA library = collection of all cDNA's from a particular cell type.) DNA from each cell type is the same; mRNA and therefore cDNA is not. See Becker fig. 23-20 (21-20).
4. If mRNA's are different, why is that? Is the difference due entirely to differences in transcription?
a. Transcription is different in different cells. It could be that all cells transcribe all genes, but only some RNA's are exported to the cytoplasm and the remaining nuclear RNAs are degraded. This is not the case. Only selected genes are transcribed in each cell type, and RNA's from those genes are processed to make mRNA.
b. Processing: Splicing and processing of same primary transcripts can be different (in different cells or at different times). Different mRNA's (& therefore proteins) can be produced from the same transcript by alternative splicing and/or poly A addition. Example below.
Try Problems 4-10 & 4-11.
IV. Regulation at
Splicing -- Results of Alternative Processing
A. There are two ways to get a collection of similar proteins
1. Gene families -- multiple, similar genes exist due to duplication and divergence of genes. Example: the globin genes constitute a family. Different family members code for myoglobin, beta-chains, alpha-chains, delta-chains, etc.
2. Alternative splicing etc (See C below) -- only one gene, but primary transcript spliced in more than one way.
B. An example of alternative splicing -- Production of antibody (immunogloblin) in B cells. See handout 11B and Becker fig. 23-31 (21-31) -- how to get either soluble or membrane-bound antibody from alternative splicing of the same transcript. (See Sadava 14.21 (14.20) for another example.)
1. Antibody can be membrane bound or secreted. Fate of antibody depends on whether peptide has a stop transfer sequence or not.
a. If has stop transfer: locks into membrane of ER and goes thru Golgi etc. to plasma membrane; stays in membrane.
b. If has no stop transfer: enters lumen of ER, goes thru Golgi etc., and then is secreted.
2. Gene has two alternative polyA addition sites. Which one is used determines final location of protein.
a. Option 1: If one (at end of exon 4/start of intron 4) is used, protein contains no hydrophobic stop transfer sequence, and protein is secreted.
b. Option 2: If other one (at end of exon 6) is used, protein contains hydrophobic sequence encoded by exons 5 & 6, and protein stays in plasma membrane.
3. mRNA can be spliced and/or poly A added in two alternate ways. Location of protein (antibody) depends on whether splicing of intron 4 or poly A addition happens first. Think of it as a competition. Either
a. Poly A adding enzymes get there before the spliceosome. In that case, poly A is added to site near end of exon 4, and rest of intron 4 (and rest of gene) is never transcribed, or
b. The spliceosome gets there first. In that case, Intron 4 is transcribed and spliced out before poly A can be added. (In this case, poly A is added at the end of exon 6 instead.)
4. Why are 2 forms of antibody needed?
a. Membrane-bound form of antibody: Serves as receptor for antigen = trap to detect when antigen is present. Binding of antigen (ligand) to antibody (receptor) serves as trigger to start secreting antibody.
b. Secreted (soluble) form: Acts as effector -- carries out major function of immune system -- binds to soluble antigen in body fluids and triggers destruction of antigen in multiple ways.
C. The general Principle -- You can get many different proteins from a single gene by the processes listed below. Therefore biologists are interested in proteomics (study of all proteins made in a cell) not only genomics (study of entire DNA or gene content).
1. Starting transcription at different points
2. Ending transcription (adding poly A) at different points
3. Splicing out different sections (exons as well as introns) of the primary transcript -- alternative splicing.
To review regulation & alternative splicing, try problems 4-12 & 4-13, & 4R-6.
V. Regulation at
translation.
A. How to control rate of translation? In principle:
1. Can regulate half life of mRNA (control rate of degradation). In prokaryotes most mRNA's have a short 1/2 life; in eukaryotes this is not necessarily so. Different mRNA's have very different half lives. (Note: proteasomes degrade only proteins NOT RNA's. )
2. Can regulate rate of initiation of translation (control how effectively translation starts).
B. Some Famous Examples of Regulation of Translation. (The principles are important; we will not go into the details.)
1. Use of a regulatory protein: -- Have a protein that binds to mRNA (or some other part of the translation apparatus) and affects either initiation and/or degradation, depending on where it binds. Two examples:
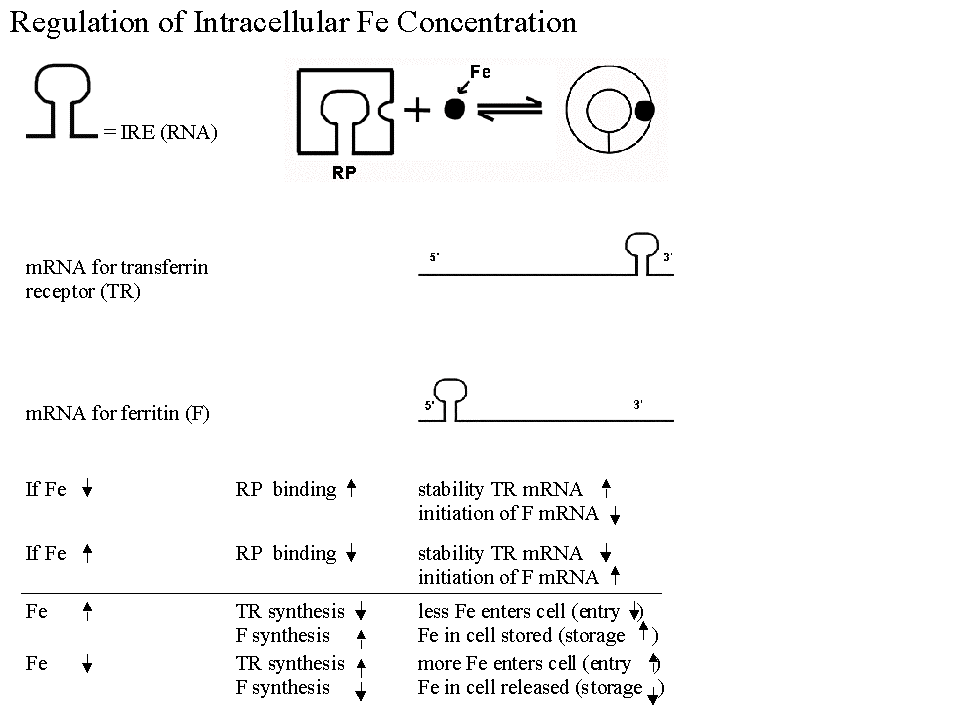
a. Regulation of synthesis of Ferritin & Transferrin Receptor (& intracellular iron levels).
- Regulatory protein acts like a (prokaryotic) repressor, but binds to regulatory sequence in mRNA, not DNA.
- Regulatory protein is allosteric, and level of small molecule effector (Fe) inactivates the regulatory protein. Note that the regulatory protein binds to mRNA in the absence of Fe, not when Fe is high. (I said it backwards in class.)
- Active form of repressor protein binds to more than one mRNA. Binds to mRNA for at protein A at 5' end (blocking initiation) and to mRNA for protein B at 3' end (blocking degradation).
- This is another example of coordinate control. There is one trans-acting factor here (repressor), but both mRNA's have the same cis acting sequence.
- See Becker, figs. 23-33 & 23-34 (21-33 & 21-34) if you are curious about the details.
A question to think about: Given the information above, which is protein A, and which is protein B? Which one is ferritin and which one is the transferrin receptor? Ferritin is an intracellular protein that stores excess iron. You can check your answer in Becker or using this diagram.
b. Regulation of globin synthesis by heme (Becker fig. 23-32). Heme (the prosthetic group of hemoglobin) stimulates synthesis of globin (the protein part of hemoglobin). In this case, heme prevents inhibition of translation.
- In the absence of heme, inhibition occurs, and translation is blocked. No globin produced.
- Heme blocks the inhibition. Therefore, in the presence of heme translation proceeds. Heme relieves the block in translation, and globin is made.
Interesting features of this case worth noting are:
(1). Inhibition of inhibition results in stimulation; in other words, (-) X (-) = (+).
(2). Another example of coordinate control. This system ensures coordination between the supply of heme and of globin. Globin is useless without heme.
(3). FYI: Heme works by interfering with phosphorylation -- it blocks a kinase from phosphorylating a critical translation factor (ElF2 = elongation factor 2). Phosphorylation of ElF2 (in the absence of heme) inhibits ElF2 and blocks translation. This is another example of a protein (ElF2) that has active and inactive forms, and phosphorylation (or dephosphorylation) interconverts the two forms. (See VI A below.)
→ Formation of double stranded RNA. This triggers degradation &/or inhibition of translation of the mRNA.2. Use of a regulatory RNA -- RNA interference (RNAi)
a. Trans acting factors can be RNA. Not all regulatory factors are protein -- some are short RNA's. (These are usually derived from double stranded RNA -- See Becker figs. 23-35 & 23-36.)
b. How does a short RNA affect translation?
(1). Inhibition (usual case): Small RNA binds to mRNA
(2). Stimulation: Some recent cases have been discovered in which small RNA binds to mRNA and 'up regulates' translation. Mechanism so far unknown.
c. Use in Regulation: Cells naturally produce micro-RNA's that bind to mRNA's and regulate translation as above. The use of short regulatory RNA's to block translation appears to be important during regulation of development. (See Becker 23-36.)
d. Use in the Lab as a tool: Called RNAi = RNA interference. The use
of artificially added short double stranded (ds) RNA to block
transcription/translation and turn genes off is very common. (See Becker
23-35.) Enzymes of cell convert added ds RNA into short single stranded
RNA that interferes with translation and/or transcription as in b. Same
effect as adding antisense RNA (but works better).
VI. Post Translational Regulation. Don't forget: regulation occurs after translation too -- after proteins are made, their activity can be modulated. Many examples of post translational modification have already come up and more will be discussed later. Here is a summary (mostly review):
A. Covalent Modification. Proteins can be modified covalently either reversibly (for ex. by phosphorylation and dephosphorylation), or permanently (for ex. by removal of N-terminal met., addition of sugars -- glycosylation, etc.)
See problem 6-3.
B. Noncovalent Modification. Proteins can be activated or inhibited by reversible noncovalent binding of other factors -- small molecule allosteric effectors, other proteins such as calmodulin (an important Ca++ binding protein to be discussed later), etc.
C. Degradation. Proteins can be selectively destroyed.
1. Half Lives Vary. Not all proteins have the same half life.
2. Proteasome: Major factor in regulation of protein turnover is control of addition of ubiquitin leading to destruction by proteasome. See Becker 23-38 (21-36) or Sadava 14.24 (14.22) or the Nobel Prizes for 2004.
3. Significance: Important example of a family of proteins that all have a short half life = cyclins; control progression through cell cycle. Different cyclins control transitions from G1 to S, G2 to M etc. Cyclins are made as needed and degraded immediately after use. (Note: Both mRNA's for cyclins and cyclins themselves are degraded after use. More on this when we cover the details of the cell cycle.)
D. Location. Proteins can activated or inhibited by a change of location. For example, transporters like GLUT4 only work if positioned in the plasma membrane; if they are sequestered in vesicles they are inactive. Transport of glucose into the cell can be regulated by moving the GLUT4 in and out of the membrane.
To review post-transcriptional &/or post-translational regulation, try problem 4-14. By now you should be able to do all the problems in 4 & 4R except for a few fine points about hormones, which should be covered below or next time.
VII. Introduction to
Signaling -- How are messages sent from one cell to another? How
are events in a multicellular organism coordinated? It's not enough to regulate what
one cell does!
A. Usual Method -- one cell secretes signal molecules that bind to a receptor on (or inside) a target cell → amplification → big effect in target cell.
B. How do secreted signal molecules work at molecular level? Overview. See handout 11A
1. Signals are evolutionarily conserved. Same signal molecules used by different organisms for different purposes.
2. Role of Receptors. First step in signaling is binding (noncovalent) of signal to receptor, causing conformational change in receptor.
3. Amplification or the Biological Big Bang. All signals → amplification = big effect from a small concentration of signal. Example: 1 molecule of epinephrine can cause release of 108 molecules of glucose from a liver cell ! (See Becker 14-3 for the calculation.)
4. How is amplification achieved? How is the 'Big Bang' Accomplished? Three ways:
a. By affecting transcription/translation → lots of new protein made → big effect
| ligand binds | → | activate a TF | → | transcribe a gene | → | make lots of mRNA molecules/gene | → | mRNA translated | → | many new protein molecules/mRNA |
Example: Thyroid hormone (also called thyrotropin or TH) & steroid hormones. Receptor is itself a TF.
b. By cascades of modification → lots of (pre-existing) protein is modified→ big effect.
General idea:
| ligand (1st messenger) binds | → | activate receptor in membrane | → | activate protein inside cell (usually a chain of activations) | → | activate target protein (enzyme, or TF, etc.) | → | lots of product |
A Specific Example: Thyroid stimulating hormone (TSH) -- promotes release of thyroid hormone (TH).
| ligand (1st messenger) binds | →* | activate receptor in membrane | → | activate G protein in the membrane | → | activate enzyme in membrane | → | generate small molecule (2nd messenger) inside cell |
Note that the ligand (1st messenger) binds to the extracellular domain of its receptor. The remaining events are inside the cell. Details of G proteins, second messengers, etc. will be discussed later. In this case 2nd messenger is cAMP, and remaining steps are:
| cAMP | →* | activate protein kinase (PKA) in cytoplasm | → | phosphorylate target enzymes | → | stimulate multiple steps in synthesis and release of TH |
Almost every step in this process involves an amplification. Steps with a * are 1:1. For all other steps, each protein modifies multiple target molecules or generates multiple molecules of product. For an estimate of the amount of amplification involved, see Becker 14-3 (target enzymes in Becker example are different, but rest is the same)**.
** The usual example for this type of modification cascade is the breakdown of
glycogen, stimulated by the hormone epinephrine (adrenaline), which is the example in Becker
fig. 14-3. If you are interested in more details of the pathway, see
Sadava 15.18 (15.15. ) or Becker fig. 14-24. (This example was the first to be
discovered, but is more complex. It will be discussed later.)
c. By opening (ligand-gated) channels
General Idea:
| ligand binds | → | open a few ligand gated channels | → | a little ion flow | → | hit threshold voltage | → | open many (voltage-gated) channels | → | big change in ion concentrations | → | big effect |
Specific example: Acetyl choline (AcCh). AcCh receptor is a Na+ channel opened by AcCh.
AcCh binds → open a few ligand gated Na+ channels → a little Na+ flows in → cell becomes less - inside; hits threshold voltage → open many (voltage-gated) Na+ channels → big change in Na+ concentrations → Muscle contracts 5. Types of Signals. Two main kinds of chemical signals -- lipid soluble and water soluble
| Signal Type | Example | Receptor Type | Effect |
| Lipid Soluble | Thyroxine, steroids | Intracellular* | Gene activity |
| Water Soluble | Peptide hormones, GF's | Cell Surface | Protein activity (usually) |
*Note: Some lipid soluble signals have cell surface receptors in addition to their intracellular receptors. One such case is in the problem book. Cell surface receptors for lipid soluble signals have been discovered relatively recently, and will largely be ignored in this course.
6. Types of Receptors -- intracellular and on cell surface See Sadava 15.4
a. Intracellular -- for lipid soluble signals. All similar, all TF's -- details later.
b. On Cell Surface -- for water soluble signals. See Becker fig. 14-2.
(1). Receptors are transmembrane proteins with an extracellular binding domain for signal. These are sometimes called "extracellular receptors" but only ligand binding domain is extracellular, not the entire protein.
(2). Three major kinds of cell surface receptors -- Listed here for reference. Details of structure/function will be discussed as we go.
(a). G Protein Linked Receptors; Also called G Protein Coupled Receptors or GPCRs. (See TSH example above. More details to follow. For an example, see Sadava 15.7)
(b). Tyrosine Kinase (TK) Linked Receptors. (These will be discussed later.) These generate cascades of modifications, but do not always use 2nd messengers. If you want to see an example, see Sadava 15.6 & 15.10 (15.6 & 15.9).
(c). Ion Channels. Receptor is part of an ion channel. (See AcCh Receptor above). To be discussed at length when we get to nerves. See Sadava 15.5.
Try problems 6-12 & 6-13.
Next time: Wrap up of whatever we don't finish. Then what are the main types of signals? How do the two basic types of signals work? What are G proteins and 2nd messengers?
{kind=link}