C2006/F2402 '08 Outline Of Lecture #2 -- Updated
01/24/08 07:27 PM
© 2008 Deborah Mowshowitz, Department of Biological Sciences, Columbia
University, New York NY
Want to see some pictures with the scanning EM? See Becker, fig. 16-5,
16A-1, 16-26, 17-1 etc.
Problems to do have been added in magenta italic bold. Problems listed below are posted on Courseworks (go to class files, and then to shared files). Problem book should be ready next week. We'll announce when it's ready. Any corrections made after lecture are marked in blue.
|
Handouts: |
1B
-- Immunofluorescence & Freeze Etch (from last time) |
I. Immunofluorescence, Antibodies, & the Cytoskeleton
A. How Immunofluorescence is Used to Visualize Cell Parts. See Becker Table 15-2 (22-2).
1. The "immuno" part: How antibodies are used as reagents to identify proteins (& other substances). See last lecture for details of a-d.
a. What are Antibodies & Antigens?
b. Specificity.
c. Antibody Structure.
d. Why use Antibodies for detection?
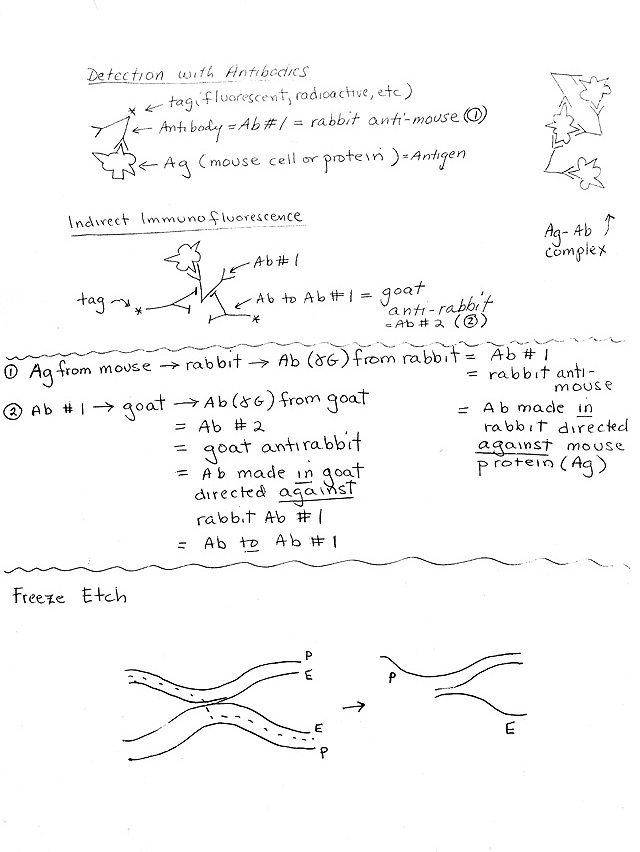
e. Detection of Ab-Ag binding. How can you tell if an antibody has bound to its antigen? Clumping and complex formation is one way. (See top of handout 1B.)
2. Role of Fluorescence See Becker Appendix, A-8 to A-11 (Guide to Microscopy p. 8-11); for pictures see Becker fig. 15-1 (22-1) or Sadava 4.3 (4.4), & 4.20 (4.21).
a. Fluorescent materials emit light of one wave length when irradiated at a different wave length.
b. Fluorescent material can be located easily by irradiating sample (at one wave length) and seeing what part of sample "lights up" (emits light at a different wave length).
c. Small fluorescent groups or small fluorescent proteins (such as green fluorescent protein or GFP) can act as "tags" or probes -- can be attached to larger molecules such as antibodies, usually to one end, often without altering function of macromolecule.
Some tags are added chemically after the protein is made by a cell.
Some tags (like GFP) are incorporated into the protein when it is made in a cell. These tags are sequences of amino acids that are added on to the normal amino acid sequence during translation. In this case, the cell must contain a recombinant gene that codes for the normal protein + the extra amino acids.
3. Methods: How Fluorescent Antibodies are used (See handout 1B)
a. General Principle: Add fluorescent antibody, wash off unattached antibodies (not bound to antigen), irradiate, and look for light emission (aka 'signal') = site of fluorescent antibody = location of target protein.
b. Direct Immunofluorescence -- Antibody with tag (fluorescent) sticks to target.
c. Indirect Immunofluorescence.-- Antibody #1 (without tag) sticks directly to target; secondary labeled (tagged) antibody sticks to constant part of first antibody. Advantages of indirect:
(1). Gives an amplification effect -- more tag or label ('signal') per molecule of target protein.
(2). Requires only one labeled antibody to identify many proteins. Same labeled secondary antibody can be used to bind to ("light up") many different proteins.
(a). A different primary antibody is used for each target protein. (Not labeled -- no tag.) Variable part of primary antibody binds to specific part of target protein.
(b). The secondary antibody binds to the constant part of the primary antibody. Therefore the same (labeled or tagged) secondary antibody can bind to many different (unlabeled) primary antibodies.
B. How Cytoskeleton Discovered
1. How do you get (labeled) antibodies to components of MT and MF? Large, permanent structures made of tubulin and actin are well known in certain specialized cells. Tubulin (from cilia & flagella) & actin (from thin filaments of muscle) purified; antibodies made to them. Fluorescent probes were attached to the antibodies, and antibodies were injected into living cells.
2. Cytoskeleton of MF & MT Identified -- During interphase, structures made of actin & tubulin found in cytoplasm of almost all eukaryotic cells.
a. Antibodies to tubulin bind to ("light up") MT.
(1). MTs are present in interphase, not just in spindle fibers during mitosis
(2). MTs are present in ordinary cells, not just in specialized cells with cilia & flagella.
b. Antibodies to actin bind to MF. MF found in ordinary cells, not just in specialized cells such as muscle.
3. Implies two possible states of tubulin and actin:
a. Temporary (dynamic) -- no fixed state; monomers and polymers are in dynamic equilibrium as in cytoskeleton, spindle fibers, cleavage furrow.
b. Permanent -- form stable specialized structures such as cilia, flagella and muscle fibers. See Sadava 4.22 (4.23) or Becker fig. 16-7 & 8 (23-7 & 23-8) for nice pictures of cilia.
4. IF found later in multicellular organisms. (Not present in unicellular eukaryotes.)
Look at problem 1-1.
II.
Microfilaments & Microtubules -- a closer look
A. Overview of Structure -- For a summary of properties, see handout 2B or Becker Table 15-1 (22-1). For pictures, see Becker fig 15-1 (22-1). For diagrams, see handout 2A (& Becker 4-24). In Sadava, see 4.20 (4.21). Many nice pictures and additional details (which go beyond the scope of this course, but might be of interest) are in Becker Chap. 15 (22).
B. How each monomer forms a polymer
→ polymer.1. Actin/MF.
a. Monomer. One type of globular monomer = G actin.
b. How polymer (filament) forms: Globular monomer (G actin) forms chain of beads. Two chains twist around each other
(1). Bind to and stabilize the polymer. Shift equilibrium (monomer ↔ polymer) to right. Ex: taxol for MT; phalloidin for MF.
(2). Bind to the monomer and prevent it from polymerizing. Shift equilibrium (monomer ↔ polymer) to left. Ex: colchicine for MT; cytochalasins for MF.
2. Tubulin/MT.
a. Monomer. Two types of globular monomers -- alpha and beta tubulin.
b. How tubule forms: Alpha + beta → dimer → chain of dimers (protofilament) → rings of chains forming a tubule (usually 13 chains/tubule).
c. Growth: Chains and tubules grow primarily by addition of dimers to "+" end.
d. Anchors:(1). Role: MT usually anchored at (and grow away from) structure at "-" end called a microtubule organizing center (MTOC) or centrosome. (For examples, see Becker Fig. 15-11 (22-10).
(2). Orientation: In most cells, MTOC and - ends are near nucleus; + ends extend toward edges of cell.
3. IF. Monomers are extended chains, not globular. Monomers are assembled into more extended chains and formed into a cable as explained below.
Look at problems 1-7 & 1-8
(A & B).
C.
Role in movement -- See Sadava fig. 4.23 (4.24)
or Becker fig. 16-3 (23-3).
1. IF not involved in movement (don't lengthen, shorten, or slide); Do assemble/disassemble during cell cycle.
2. MF, MT involved in movement. Primarily using a "motor molecule" (see table on handout) that slides or 'walks' down a fiber using the energy from splitting of ATP.
a. Two major motor molecules for tubulin/MT:
(1) Dynein -- Dynein carries material "in" toward the nucleus/cell body -- Dynein draws things in (toward the - end of MT).
(2) Kinesin -- Kinesin carries material "out" toward the edges of the cell -- to the cell "Korners" (toward the + end of MT).
b. One major motor molecule for actin/MF = myosin
3. Two major types of movement using motor molecules
a. Can have 2 fibers sliding past each other (motor is part of one fiber or in between the two)-- overall effect is to shorten/lengthen structure. See Sadava fig. 4-23 (a) (4-24 (a)). Examples:
(1). Anaphase -- MT slide "out" to give longer spindle fibers and elongated spindle.
Note: Overall movement of chromosomes seems to depend both on sliding of some fibers past each other, and the change in length of other individual fibers. For details, see Becker.
(2). Telophase -- MF slide "in" forming cleavage furrow that divides cell in half. Muscles contract when MF slide in.
b. Can have vesicle or large structure moving down a fiber (motor attached to vesicle) -- fiber acts as "railroad tracks" to direct vesicle toward one end of fiber. Example -- How vesicles move down axons of neurons. See Becker fig. 16-1, 16-2 & 16-5 (13-1, 23-2 & 23-5) or Sadava 4.23 (b)& (c) (4.24 (b) & (c)).
(1). Structure: Neurons are nerve cells with long extensions (axons) that make connections (synapses) with other cells at the end of the axon. Multiple MT extend through the length of the axon.
(2). Vesicle Transport: Vesicles are transported along the MT between nucleus/cell body at near end of axon and synapse at far end of axon. Direction of movement depends on whether motor molecule is dynein or kinesin.
(3). Important Note: electrical signals are not transmitted down axons using vesicles -- vesicle transport and signal transport are two separate processes. Electrical signal transport is much faster, and will be explained later.
For an animation of a vesicle moving down a fiber (& many other fascinating cell features) see the video from Harvard called "The inner life of the cell." A shorter version of this video (set to music) is available on many web sites including http://www.youtube.com/watch?v=CVUnzk40npw and http://multimedia.mcb.harvard.edu/media.html. The Harvard web site also has a longer version with narration instead of the music. Many of the things shown in the video may be obscure at this point, but by the end of the cell biology section of this term you should be able to identify almost all the items shown.
Look at problems 1-9 to
1-11. See handout 2-B for effects of drugs mentioned.
III. IF's
-- see handout 2A
A. How monomers form a polymer
1. Monomers: All Monomers are extended, not globular -- see handout. There is more than one type of monomer (see below).
2. Formation of polymer:
a. Two monomers → dimer; both monomers point in the same direction. Two monomers may be same (homodimer) or different (heterodimer).
b. 2 dimers → tetramer; tetramer = fibrous basic subunit. Two dimers in a tetramer point in opposite directions.
c. Tetramers stick to each other by overlapping ends → protofilament.
d. Multiple protofilaments form a cable, probably 8 protofilaments across. Exact final structure unclear. One possibility: protofilaments form flat cable that twists into final structure. (fig. 15-25(22-25) of Becker = handout 2A.)
Note: Becker text says tetramer = protofilament; this appears to be an error. All other texts and Becker diagram describe protofilament as a long structure formed by many tetramers.
B. Not all IF's are the same. All IF's have similarities, and form similar structures, as above, but there are multiple IF genes and therefore multiple IF's with significant differences. (See Becker fig. 15-24 (22-24) = handout 2A)
1. Nuclear IF's -- Lamins. Lamins are a type of IF found in the nucleus. Do not confuse them with laminins (proteins found in the extracellular matrix). Lamins are the same in all cell types.
2. Cytoplasmic IF's
a. All IF's but lamins are cytoplasmic. (All MF & MT are cytoplasmic.)
b. Cytoplasmic IF's (not lamins) are tissue specific -- origins of cancers can be traced from the type of IF's they contain. That's because different tissues transcribe ("express") different IF genes.
C. IF's represent a protein/gene family. See Sadava Ch. 24 esp 24.2 (Ch 26 esp. pp. 516-519) for more on evolution of gene families.
1. Proteins tend to occur in "families" -- groups of similar proteins.
2. Examples: all IF's (& their genes) are similar. All globins (Hb alpha chains, Hb beta chains, myoglobin) are similar to each other but very different from all IF's. All antibody chains are similar to each other but not to globins and IF's, and so on.
3. How do families form? All members of a family have a common evolutionary origin -- ancestral gene duplicated and copies diverged → family of related proteins.
a. Why do copies stay so similar? Sections of the protein (& corresponding sections of the gene) that were essential to IF formation were preserved in all duplicates -- mutations that caused loss or serious alteration of these sections were lost (= conservative selection against mutations that ruin function = selection against organisms that function worse than average).
b. Why do copies diverge? Alterations that allowed useful variations in function were preserved (= innovative selection for mutations that improve function = selection for organisms that function better than average).
4. Two ways to 'fine tune' protein structure & get a group of similar, but different, proteins: By gene duplication to give a gene family OR alternative splicing of a single gene.
IV. Introduction to Membrane Structure This will be covered next time.
A. Lipid part
1. Amphipathic nature of lipids -- See Sadava fig. 5.2 -- there are multiple different "two headed" lipids -- each type has a different structure, but each has a hydrophobic end and hydrophilic end.
2. Amphipathic Lipids form a bilayer.
B. Protein part -- where are the proteins (relative to the lipid)? Is it a "unit membrane" or a "fluid mosaic?"
For "unit membrane" See Becker fig. 7-4 ; for fluid mosaic model see Becker fig. 7-5 (or 7-3) or Sadava 5.1.
1. Use of freeze fracture/freeze etch procedure
a. E vs P faces of bilayer = surfaces you see if you crack bilayer open = inside of bilayer
(1). E face = inside of the monolayer that is closer to extracellular space (outside of cell)
(2). P face = inside of the monolayer that is closer to protoplasm (inside of cell)
b. What do you see on inside? (see Becker fig. 7-16 & 7-17 or Sadava 5.3).
(1) Inside is not smooth -- shows proteins go through bilayer (implies "mosaic" model not unit membrane)
(2). More bumps (proteins) on P face than E face -- shows more proteins anchored on cytoplasmic (protoplasmic) side.
c. Freeze fracture vs Freeze etch
(1). Freeze fracture = crack frozen sample open, examine in EM;
(2). Freeze etch = crack open, let some water sublime off, then look in EM. For some sample pictures, see Becker figs. 15-16, 15-21, 15-26, & 16-1. (22-19, 22-21, 22-26, & 23-1.)
C. Fluid mosaic model -- overview of current idea of how proteins and lipids are arranged. See Becker fig. 7-5 (or 7-3) or Sadava 5.1.
Next Time: Details of membrane structure; then on to transport -- how are small molecules moved across membranes?
{kind=link}