C2006/F2402 '09 -- Lecture 1 -- Updated 01/26/09
© 2009 Deborah Mowshowitz, Department of Biological Sciences, Columbia University, New York NY
Handouts: 1A -- Eukaryotic
Cell Structure ; 1B --
Immunofluorescence;
1C -- Overview
of Course (Brief Description)
There are additional handouts from Science Magazine. These
are from a special section on breakthroughs in '08 and predictions for '09. An
expanded version of the special section is available online at:
http://www.sciencemag.org/btoy2008/
Class handouts 1A and 1B are slightly different from the ones on line. Copies of all class handouts and returned exams are available on the 7th floor of Mudd, outside Dr. M's office.
Class handout titled 'Overview' is not comprehensive. Be sure to check out 'About C2006' or 'About F2402.'
References to Becker are to 7th ed. (When fig. numbers or pages in 6th edition are different, refs. to 5th ed. follow in parentheses.) Refs to Sadava are to 8th ed of Life (7th in parentheses).
The online notes for this term will be extensive outlines, not word-for-word. This is partially for practical reasons and partially for educational reasons -- I think it encourages better student note taking and understanding. The outlines will be provided the night before (if not earlier) so you can print them out in advance and annotate them in class. If you like to read ahead, the outlines from last year (2008) are linked to the '08 schedule. Older schedules, with references to readings in older editions of the texts, are available too, on the old schedules page.
I. The
Story So Far -- Summary of last Term
A. The Major Question: How do Living Things Work? Or how does 1 cell make two?
B. Answer so far has concentrated on
1. Prokaryotes (mostly) -- 1 intracellular compartment, unicellular
2. Macromolecular level -- did not discuss larger structures such as organelles, chromosomes, etc.
3. The "big 5" issues -- structure, function, manufacture (including energy requirements), regulation, and (evolutionary) origin.
4. Methods -- those used to figure out how living things work, especially the methods used to get at the "big 5."
C. Bottom Line: DNA
→ RNA
(& more DNA); RNA →
protein → job, say catalyzing X → Y. Protein production & activity
are
regulated.
We emphasized roles of mRNA, tRNA & rRNA in translation, which are well
known. Roles of short regulatory RNAs (micro RNA's , RNAi) in regulation of translation &
transcription were mentioned only briefly, as details are just now being uncovered.
See 'Scorecard' on handout 1D. Micro RNA was rated one of the 'Areas to Watch' in Dec. 2007.
II. Overview of this Term
A. Similarities: Same Major Question; Same "big-5" issues; same stress on methods.
B. Differences:
1. Organisms: This term will concentrate on Eukaryotes (mostly) as vs prokaryotes. See Becker table 4-1 for comparison of Eukaryotic and Prokaryotic cells. For pictures, compare Sadave 4.4 (4.5) & 4.7.
2. Structures: This term will consider many things bigger than macromolecules -- intracellular structures (organelles, etc.) and assemblies of cells (organs, tissues, stystems).
Last Term This Term Organisms Prokaryotes Eukaryotes Structures Macromolecules Bigger
3.
New Issues: Two big innovations of eukaryotic cells are:
a. Intracellular Compartmentalization
b. Multicellularity
4. Topics: Most of the topics this term will deal with the consequences of the two big innovations.
Last Term This Term Biochemistry Cell Biology Genetics Signaling Molecular Biology Development Physiology
C. Major Subjects of Term
1. Cell Bio -- study of consequences/implications of compartmentalization within cells (Lectures 1-10)
a. Macromolecules and bigger things (membranes and organelles -- chromosomes, etc.) Lectures 1-3.
b. Co-ordination (between different parts of cell) -- how do things get to the right compartment? How do they cross membranes? Lectures 4-8.
c. Regulation of eukaryotic gene expression -- how do things get made at the right time (as well as in the right place)? Lectures 9 & 10.
d. Note: in this part, we will emphasize generalized features of all eukaryotic cells but discuss some specialized cell types.
2. Signaling between cells & some of the consequences -- the need for co-ordination between different cells in multicellular organisms.(as vs co-ordination between the different parts of a single cell). Some examples of how specialized cells carry out specialized functions.
a. How cells communicate chemically (Lectures 11-12). Any aspects of this that are not finished will be wrapped up in lecture 19, along with how cell-cell communication controls the cell cycle.
b. How cell communicate electrically ( Lectures 13 & 14 by Dr. Stuart Firestein; structure and function of nerve cells.
c. Wrap up of nerves and integration with Muscle (Lectures 15 & 16).
3. Development -- How
do you build a multicellular organism? How do cells get specialized and how they stay
that way? Lectures 17 & 18 by Dr. Alice Heicklen.
For a taste of what's to come, see the handout on 'Reprograming Cells' that
Science magazine describes as the #1 'Breakthrough of the Year' for '08. For
an expanded version of the print magazine special section, go to
http://www.sciencemag.org/btoy2008/
4. Physiology -- Study of selected consequences/implications of multicellularity for maintenance of organism as a whole. How do you run a multicellular organism, once you've built it? How organisms make use of communication, specialization, etc. to maintain a relatively constant internal environment and how all the various functions are co-ordinated. This subject will be divided as follows:
a. Homeostasis & Hormones: How the various systems (nervous, hormonal, muscular, respiratory, etc.) work to maintain a constant internal environment of temperature, gas, fluid, salt & water (Lectures 20-23) and respond appropriately to changes in external environment.
b. Immunology -- The most specialized system; how a multicellular organism fends off invaders (Lecture 24).
5. Cell cycle & Cancer: How aberrations in cell-cell communication and/or the cell cycle may cause cancer (Lectures 25).
III. Eukaryotes -- a closer look.
B. Issues of resolution -- For pictures using the different methods of microscopy, see Becker table 1-1 or Sadava 4.3 (4.4.) For more details on microscopy, see Becker appendix.
Types of Microscopes -- Light microscope, SEM (scanning electron microscope) and TEM (transmission electron microscope)
|
Limit of Resolution |
Useful Magnification |
Features |
|
|
Eye |
0.1 mm |
1 |
|
|
Light Microscope |
0.2 micron |
1000X |
Can look at living specimens |
|
TEM |
1-2 nm (in practice) |
100,000X or 100X light scope |
Detect electrons from source that pass through thin slice of tissue |
|
SEM |
Larger than TEM but gives 3D picture of surface* |
Less than TEM |
Detect secondary electrons emitted from surface of sample |
* For a nice SEM picture see Becker fig. 1-3 (1-3).
For more SEM pictures of biological
specimens see
http://www.le.ac.uk/bs/em/sem.htm.
For a completely different use of the scanning EM see
http://www.tfhrc.gov/pubrds/05may/03.htm.
Sizes of things commonly encountered in biology
|
1/1000 |
1/1000 |
1/1000 |
1/10 |
|||||
|
m (meter) |
→ |
mm |
→ |
micron (μ) |
→ |
nm (nano) |
→ |
Angstrom (Å) |
mm = millimeter = 10-3 meter
micron (μ) = micrometer or 'mu' = 10-6 meter
nm = nanometer = 10-9 meter
Angstrom (Å) = 10-10 meter
Sizes of some cells, structures and very small living (?) things:
| Cell Type | Diameter |
| Typical eukaryotic animal cell | 50 microns (μ) = 50,000 nm. |
| Typical bacterium | 1-5μ = 1-5000 nm |
| Smallest known bacterium | 0.2 μ = 200 nm |
| nanobe (for Times article see; for more info see) | 20-150 nm |
| ribosome | 25-50 nm |
For more examples and tables of relative sizes see Becker fig.1-3 (1-2) & box 1A, or Sadava 4.1 (4.2) .
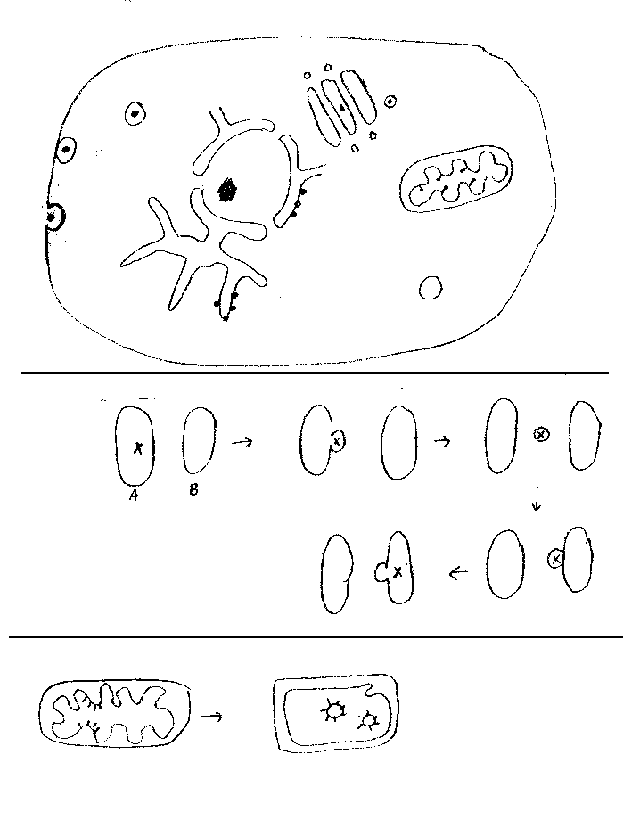
C. What you can see in the EM? See handout 1A and Becker fig. 4-5 or Sadava 4.7 for pictures of whole euk. cells. Some specific references to pictures of organelles are given below, but all pictures in Ch. 4 of both texts are well worth it. Methods used to characterize the various parts are described below in IV, but will be discussed as they come up.
1. Membranes. In the EM you can actually see membranes (6-9 nm across); Membrane = bilayer of lipid plus associated proteins. In the EM, after staining, bilayer appears as a double line. (Can also be described as a three layered structure, with two dark layers sandwiching a lighter layer between them. Only the top and bottom layers are visible in the EM = double line.) Detailed structure of membranes will be covered next time. Note that a bilayer = a single membrane. A 'double membrane' = two complete bilayers. (See nucleus, below.)
2. Nuclear details -- See figs. 4-4 & 4-10 of Becker or 4.8 (4.9) of Sadava or go to Google images and search.
Nucleolus -- not membrane bounded
Nuclear Pores & Nuclear Envelope (Double membrane = 2 bilayers, with holes punched across both layers)
Space between two layers of nuclear envelope = Perinuclear Space
No whole ribosomes in the nucleus. (Does contain newly made subunits on their way 'out' to the cytoplasm.)*
* The majority opinion is that there is are no whole ribosomes and no translation in the nucleus. However there is some evidence that has been interpreted to indicate the presence of a few whole ribosomes carrying out translation in the nucleus.
3. Endomembrane System -- See Sadava figs. 4.10-4.12 (4.11 to 4.13) or Becker fig. 4-15 to 4-18) For a more detailed picture, see fig. 12-1 of Becker.
a. Components
ER (smooth & rough) -- continuous, but only rough (RER) has attached ribosomes. Membranes of ER are continuous with outer layer of nuclear envelope.
Golgi
Many types of Vesicles. (Some examples: lysosomes, transport vesicles, secretory vesicles, & endosomes.) Name, structure & function of each type will be discussed later -- features of each type depend on role in processing, packaging, transport, etc.
b. Major function = processing, sorting, packaging and transport of proteins to/from outside of cell or proper part of EMS.
c. Structure of Components
All parts are made of vesicles and flattened sacs (cisternae). Shapes vary.
Each vesicle or sac surrounded by a single membrane (one bilayer)
Inside of each vesicle/sac = lumen = space inside a hollow organelle, organ, or tube.
Why it's considered one system: Material can be transferred from one lumen to another -- therefore all lumens effectively (but not physically) connected -- see below.
Important: Not all vesicles are part of the EMS.
d. Isolation: See methods section below for how
the various components are isolated and/or distinguished from each other.
e. Exocytosis and endocytosis--
Unique to eukaryotic cells. See
handout 1A for steps
All internal spaces of endomembrane system (lumens & perinuclear space, but not inside of nucleus) and the outside of the cell are effectively connected -- material can be passed between any of these spaces and the outside of the cell.
How are these spaces & outside of cell (effectively) connected? Through formation and fusion of vesicles.
Vesicular traffic (budding off and fusion of vesicles) moves and carries both "cargo" (vesicle contents) and membranes.
Vesicle traffic can transport cargo two ways (see handout 1A).
(a) between one membrane-bound compartment inside the cell to another or
(b) between inside and outside of cell (by exo- & endocytosis).You need labeling of cargo to determine direction.
4. Peroxisomes (See Becker fig. 4-19 & 4-20 or Sadava 4.17 (4.19)
a. Peroxisomes & lysosomes have some common features:
Are surrounded by a single membrane (unlike nucleus or endosymbionts).
Contains enzymes in lumen
Can be about the same size
Contain reactive materials that can destroy cytoplasmic molecules
b. A critical difference: Different enzymes are found inside the 2 organelles
Lysosomes contain acid hydrolases
Peroxisomes contain oxidases
Note: The two organelles arise from different structures as will be explained later. Only lysosomes are part of the EMS.
c. Are peroxisomes & lysosomes 2 different organelles? How do we know? See methods below.
5. Mitochondria and chloroplasts
a. Structures -- see texts (Becker fig. 4-11 & 4-14; Sadava 4-14 & 4-14 (4-14 & 4-15.) Note double membranes (2 bilayers), prokaryotic style ribosomes, circular DNA, overall size similar to that of bacteria.
b. Have own genetic systems (DNA, ribosomes etc. -- everything needed for DNA replication, transcription & translation.) Mitochondrial DNA is often used for identifications (when nuclear DNA is not available) or for tracing inheritance of the female line. Peroxisomes, lysosomes etc. do NOT contain DNA, ribosomes, etc.
c. Most proteins of mito and chloro are NOT made inside the organelle -- most are encoded in the nucleus, made in the cytoplasm, and transported into the organelle after synthesis.
d. There are strong similarities between mito &. chloro -- electron transport in membrane used to pump protons, H+ gradient used to make ATP; carbon rearrangements carried out in matrix or equivalent, etc. Exergonic process that drives electron transport is different -- light absorption (in chloro) vs oxidation of reduced carbon compounds (in mito).
e. Origins -- probably endosymbionts. Mito. and chloro. were once probably free living bacteria. See Sadava 27.7 (4.18 & 28.3) or Becker Box 11A.
Q's to keep in mind: (1) What are the major similarities and differences between peroxisomes, lysosomes and mitochondria? How can you tell them apart? It is a good idea to make a table that compares and contrasts the three organelles. (2) How are new organelles made? The answers are different for the 3 types of organelles and will be discussed in detail in a few lectures.
6. Location/function of ribosomes.
Two types of ribosomes are making protein in cytoplasm -- "free" ribosomes or ribosomes attached to membranes of ER.
Free cytoplasmic ribosomes (attached to mRNA, but not to ER) make proteins for nucleus, cytoplasm, peroxisomes etc. -- all parts of cell except EMS.
Bound ribosomes on RER make proteins for EMS or outside the cell.
How the "right" ribosomes attach to the ER & how the various proteins reach their correct destinations will be discussed at length in lectures 6-8.
Additional ribosomes (prokaryotic in size and function) are found inside mitochondria and chloroplasts.
7. Cytoskeleton (see Becker figs. 4-23 & 4-24 or Sadava 4.20 (4.21) -- we will discuss this in more detail next time.
a. Three components made of protein
| Component | abbreviation | shape | Made of: | Diameter |
| Microtubules | MT | hollow | tubulin | 25 nm |
| Microfilaments | MF | solid rod | actin | 5-9 nm |
| Intermediate filaments | IF | cable | Different proteins; all similar (in same family); Found in cells of multicellular organisms only |
8-12 nm |
b. How discovered? By immunofluorescence (see below) using antibodies to tubulin and/or actin.
c. Functions -- more on structure & function next time
a. Support/strength -- weight bearing, shape determining.
b. Movement (MF & MT)-- can change shape of cell (as in muscle or amoeba) and/or help move organelles within cells.
c. Localization of other factors -- act as peg board or framework for attachment of organelles, enzymes, etc. (Cytoplasm is not simply a "bag of enzymes.")
IV. Methods -- these will be discussed in Lecture #1 or #2 as we get to them.
How do you find out which components are in each cell part?
Reminder:
Becker has a guide to all techniques and methods described in the book -- see
inside front cover.
A. "Grind and Find" (Biochemical separations and assays). See Becker Box 4B and Box12A or Sadava 4.6 (4.8).
1. Grind: Break up cell into parts, and separate (fractionate) parts by ultracentrifugation.
2. Find: Find the cell part associated with each particular enzyme. How? Assay (test) each cell fraction for enzymes of interest. (Provide substrate for enzyme; look for disappearance of substrate or formation of product.)
B. In situ labeling (Localization of enzymes "in situ" = in place).
1. Substrate: Provide solution of substrate. Enzyme substrate is soluble, so it diffuses to site of enzyme.
2. Product: Product of enzyme catalyzed reaction is insoluble, so it stays in the place where it is made. If cells are washed, unused substrate will be removed, but product will stay put.
3. Localization: Insoluble product is produced and precipitates only at location of enzyme -- pin points position of enzyme.
4. Detection of product: Can be colored (for detection in light microscope) or electron dense (for detection in EM). For an example, see Becker fig. 12-20.
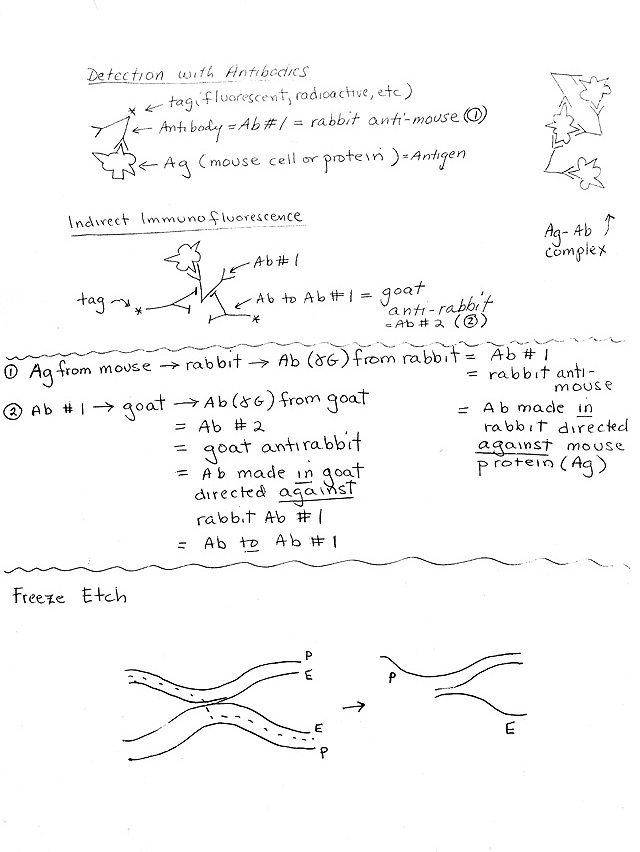
C. Immunofluorescence -- Use of labeled antibodies as tags -- direct (one step) and indirect (two step). Handout 1B. See Becker table 15-2. For more details, see Appendix, A-8 to A-11. For a picture of a typical result, see Becker table 15-1 or Sadava 4.3 (4.4). Allows you to visualize location(s) of particular proteins.
1. The "immuno" part: How antibodies are used as reagents to identify proteins (& other substances)
a. What are Antibodies & Antigens? Antibodies are made by vertebrates in response to foreign materials (antigens). Antibodies are always proteins; antigens can be proteins (as in all cases discussed here) or other substances.
b. Specificity. Each antibody (against a protein) binds to one protein or a very small number of similar proteins. (See Becker fig. 15-10 for an example with a metal labeled antibody; table 15-1 for examples with fluorescently labeled antibodies.)
c. Antibody Structure.
(1). Each antibody has a variable part -- complementary in fit to small part of target (in this case, protein.)
(2). Each antibody has a constant part -- constant in all antibodies of that class from that species.
d. Why use Antibodies for detection? Many methods identify (or characterize) proteins by their function; antibodies identify proteins by their structure (irrespective of function). Therefore antibodies are often useful for detection of proteins that have no enzymatic activity (such as components of cytoskeleton).
e. Detection of Ab-Ag binding. How can you tell if an antibody has bound to its antigen? Clumping and complex formation is one way.
2. Role of Fluorescence See Becker Appendix, A-8 to A-11; for pictures see Becker table 15-1 or Sadava 4.3 (4.4), 4.21 (4.20).
a. Fluorescent materials emit light of one wave length when irradiated at a different wave length.
b. Fluorescent material can be located easily by irradiating sample (at one wave length) and seeing what part of sample "lights up" (emits light at a different wave length).
c. Small fluorescent groups or small fluorescent proteins (such as green fluorescent protein or GFP) can act as "tags" or probes -- can be attached to larger molecules such as antibodies, often without altering function of macromolecule.
Some tags are added chemically after the protein is made by a cell.
Some tags (like GFP) are incorporated into the protein when it is made in a cell. These tags are sequences of amino acids that are added on to the normal amino acid sequence during translation. In this case, the cell must contain a recombinant gene that codes for the normal protein + the extra amino acids.
The 2008 Nobel Prize in Chemistry was awarded for the discovery and development of the use of GFP. One of the awardees was Dr. Martin Chalfie, chair of the Dept. of Biological Sciences at Columbia. For the presentation speech (describing the significance of GFP) go to
http://nobelprize.org/nobel_prizes/chemistry/laureates/2008/presentation-speech.html.3. How Fluorescent Antibodies are used (See handout 1B.)
a. General Principle: Add fluorescent antibody, wash off unattached antibodies (not bound to antigen), irradiate, and look for light emission = site of fluorescent antibody (= site of target protein).
b. Direct Immunofluorescence -- Antibody with tag (fluorescent) sticks to target.
c. Indirect Immunofluorescence.-- Antibody #1 (without tag) sticks directly to target; secondary labeled (tagged) antibody sticks to constant part of first antibody. Advantages of indirect:
(1). Gives an amplification effect -- more tag or label ('signal') per molecule of target protein.
(2). Requires only one labeled antibody to identify many proteins. Same labeled secondary antibody can be used to bind to ("light up") many different proteins.
(a). A different primary antibody is used for each target protein. (Not labeled -- no tag.) Variable part of primary antibody binds to specific part of target protein.
(b). The secondary antibody binds to the constant part of the primary antibody. Therefore the same (labeled or tagged) secondary antibody can bind to many different (unlabeled) primary antibodies.
Next time: More on the cytoskeleton, and the structure of cell membranes.
{kind=link}
{kind=link}