C2006/F2402 '09 OUTLINE OF LECTURE #11
(c) 2009 Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last update 02/26/2009 02:17 PM .
Handouts: Handouts are 11A -- RNA processing; 11B -- Alternative Processing/Splicing
I. Overall Regulation of Eukaryotic Gene Expression -- What has to be done to make more or less of a protein? A different protein? What steps can be regulated?
A. If cells make different proteins, how is that controlled? If two eukaryotic cells (from a multicellular organism) make different proteins, what is (usually) different between them?
Examples: Chicken oviduct makes ovalbumin -- chicken RBC make globin
Human liver makes transferrin -- human precursors to RBC make globin
1. Is DNA different? (No, except in cells of immune system.)
2. Is mRNA different? (Ans: yes). This means you can get tissue specific sequences from a cDNA library. (cDNA library = collection of all cDNA's from a particular cell type.) DNA from each cell type is the same; mRNA and therefore cDNA is not. See Becker fig. 23-20.
3. Is state of chromatin usually different? (Ans: yes) How is this tested? Method & result described previously. See figure 23-17 in Becker.
4. Why is the state of the chromatin different? Is the difference in the cis acting regulatory sequences or the trans acting factors? (Answer to be discussed in class.) See Becker fig. 23-24.
5. If mRNA's are different, why is that? Is the difference due entirely to differences in transcription?
a. Transcription is different in different cells. It could be that all cells transcribe all genes, but only some RNA's are exported to the cytoplasm and the remaining nuclear RNAs are degraded. This is not the case. Only selected genes are transcribed in each cell type, and RNA's from those genes are processed to make mRNA. (For an experiment that shows this, see figure 23-19 in Becker.)
b. Processing: Splicing and processing of same primary transcripts can be different (in different cells or at different times). Different mRNA's (& therefore proteins) can be produced from the same transcript by alternative splicing and/or poly A addition. Details & example below.
To review transcription, try problems 4R-5 and 4R-6A.
B. How can amount of protein synthesized be controlled? If cell makes more or less protein, which step(s) are regulated?
1. In prokaryote (for comparison) -- process relatively simple.
a. Most regulation at transcription.
b. Translation in same compartment as transcription; translation follows automatically.
c. Most mRNA has short half-life.
2. In eukaryote -- Gene expression has many more steps & complications than in prokaryotes -- more additional points of regulation -- not just at transcription. See Becker fig. 23-11 or Sadava 14.12 (14.11).
a. Transcription is main point of control, but other steps are often regulated too.
b. Transcription & translation occur in separate compartments. Translation does not follow automatically.
(1). 2. Transcript must be processed (capped, spliced, polyadenylated, etc.) -- any of these steps can be regulated, and there is more than one way to process most primary transcripts.
(2). mRNA must be transported to cytoplasm.
(3). Translation can be regulated (independently of transcription) -- can control usage and/or fate of mRNA, not just supply of mRNA. For any particular mRNA, can regulate 1 or both of following:
(a). Rate of initiation -- can control how often ribosomes attach and start translation.
(b). Rate of degradation -- can control half life of mRNA.
c. Different eukaryotic mRNAs have different half-lives. Some mRNA's are long lived and some have a very short half life.
To review regulation, try Problems 4-11 & 4-12.
II. Processing of Eukaryotic mRNA transcripts
A. Caps and poly A -- See handout 11A.
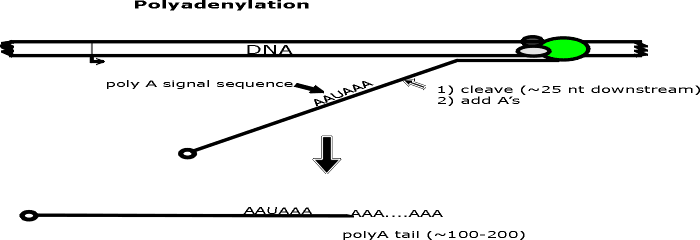
Most eukaryotic transcripts that will be used as mRNA must be modified on both ends (as well as spliced) before they can be transported to the cytoplasm and used for translation. A "cap" is usually added to the 5' end and a "poly A tail" to the 3' end. The steps involved generally precede splicing and are shown on handout 11A. (Numbers below match steps on handout.)
(1) Beginning of transcription.
a. The start of transcription is usually indicated by a bent arrow.
b. The boxed areas of the DNA = exons; plain DNA between them = intron.
(2). Capping.
a. A modified G is added to the 5' end of the transcript shortly after transcription begins, while the transcript is still being made.
b. The G is added "backwards" so there is a 5' to 5' connection. (For the curious: The structure of the cap and how it is connected to the transcript is shown in your texts; see fig. Becker 21-18.)
c. The cap is represented on the handout as a filled circle.
(3) Transcription continues to or slightly beyond the end of the gene or transcription unit.
a. There may be no fixed stop for transcription in eukaryotes (for production of most mRNA); the addition of poly A (see below) may determine the exact 3' end of the transcript.
b. Most but not all eukaryote mRNA's contain poly A.
c. Reminder: in eukaryotes production of rRNA & tRNA are carried out by different RNA polymerases which have somewhat different properties. these RNA's do not have poly A. (For details see texts.)
(4 & 5). Polyadenylation.
a. A poly A tail -- a string of A's a few hundred long -- is added to the 3' end of the RNA.
b. Growth of AA...... is 5' to 3' using ATP and splitting off pyrophosphate as usual. No template is used.
c. The sequence AAUAAA is the signal for the appropriate enzyme to cut the transcript a bit downstream and add the string of A's. (Downstream = in the 3' direction on the mRNA or sense strand.)
d. Note that the A's on the 3' end and the G of the cap are not encoded in the template DNA.
e. On the handout cleavage of transcript = step 4; addition of poly A = step 5. These two steps may occur simultaneously.
(6) Set up for Splicing. Nothing has happened to the RNA in step 6 except that it has been labeled to indicate exons and introns (so we can explain splicing).
a. By the time the transcript is released from the DNA it already has a cap on the 5' end and a poly A tail on the 3' end. This RNA -- modified on both ends but not spliced -- is usually called the primary transcript or pre-mRNA.
b. Some texts refer to unmodified RNA as the primary transcript, but such a state doesn't really exist, since the pre-mRNA is modified before it is released from the DNA.
c. The RNA is now ready for splicing (steps 7-9 on handout). See also Becker fig. 21-22 (21-23) or Sadava 14.10 (14.9).
B. Splicing of Eukaryotic mRNA
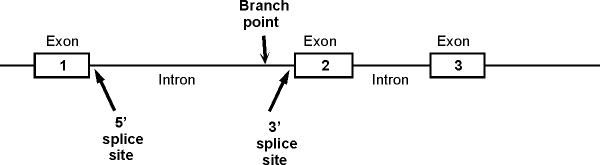
1. A typical picture of a gene with introns and exons (for reference). The picture below shows a section of the sense strand of the DNA that includes a gene with 3 exons and 2 introns. (The picture on the handout has 2 exons and one intron.) Conventions:
The picture on the handout shows double stranded DNA, but genes are often shown as in the picture below, with only the sense strand actually drawn in.
Transcription would start at the 5' (left) end of exon 1 and go to the right.
Important features of intron: Branch point, 5' splice site (also called the donor site) and 3' splice site (also called acceptor site). These are shown for the first intron only. (See also fig. 21-22 (21-23) in Becker or 14.11 (14.10) in Sadava.)
Also note that the region to the left of exon 1 is NOT an intron -- it is not part of the gene. It is part of a spacer in between this gene and the previous one.
2.
Splicing Details -- See bottom of handout 11A.
a. General Features
(1). Splicing out of each intron occurs in 3 steps (see handout 11A, steps 7-9). At each step the parts of the transcript are held in place by the spliceosome. The steps are repeated for splicing of each intron -- many RNA's have many introns. Details are below.
(2). The splice junction at the 5' end of an intron is called the 5' or donor site; the splice junction at the 3' end of an intron is called the 3' or acceptor site.
b. Steps of splicing. See handout 11A at bottom. See also Becker fig. 21-24 or Sadava 14.11 (14.10). All the steps are catalyzed by the spliceosome. Steps on handout are as follows:
(7) RNA transcript forms loop for removal of intron.
(8). Cut at 5' end of intron
(a). 5' splice site (donor site) is cleaved
(b). loose end of intron (5' end of intron) attaches to branch point in the middle of the intron, forming lariat-shaped structure.
(9). Final Step
(a). The 5' donor site attaches to the 3' acceptor site, joining the two exons and releasing the intron in the form of lariat.
(b). The lariat will be degraded and the nucleotides will be recycled.
(c). The RNA containing the exons (without the introns) will be transported to the cytoplasm and translated.
c. N.B: Prokaryotes do not have introns and lack the machinery needed to remove them.
3. Do exons and translated regions coincide? See diagram at bottom of 11A.
a. Exons include UTR's. The start and stop signals for translation are not at the
ends of the exons -- the exons include untranslated regions.
(1). Leaders. At the 5' end, there is a 5' untranslated region (UTR) or
leader before translation begins (before the first AUG) that is part of the
first exon.
(2). Trailers. At the 3' end, there is a 3' UTR
or trailer that is after the stop codon that is part of the last
b. Therefore the exons are not = protein coding sequences, as some texts imply. (The diagram in Sadava 14.5 (14.4) is incorrect.)
c. Exons = sections of genes that are represented in the mRNA. Exons include untranslated 5' and 3' regions as well as the translated regions.
d. Is an exon a "coding region"? It is a "coding region" in the sense that it codes for mRNA, but NOT in the sense that it (necessarily) codes for protein. Exons include sequences that are represented in the mRNA but do not code for amino acids.
III. Regulation at
Splicing -- Results of Alternative Processing
A. There are two ways to get a collection of similar proteins
1. Gene families -- multiple, similar genes exist due to duplication and divergence of genes. Example: the globin genes constitute a family. Different family members code for myoglobin, beta-chains, alpha-chains, delta-chains, etc.
2. Alternative splicing or processing (See C below) -- only one gene, but primary transcript spliced in more than one way.
B. An example of alternative processing -- Production of antibody (immunogloblin) in B cells. See handout 11B and Becker fig. 23-31 -- how to get either soluble or membrane-bound antibody from alternative processing of the same transcript. (See Sadava 14.21 (14.20) for another example.)
1. Antibody can be membrane bound or secreted. Fate of antibody depends on whether peptide has a stop transfer sequence or not.
a. If has stop transfer: locks into membrane of ER and goes thru Golgi etc. to plasma membrane; stays in membrane.
b. If has no stop transfer: enters lumen of ER, goes thru Golgi etc., and then is secreted.
2. Gene has two alternative polyA addition sites. Which one is used determines final location of protein.
a. Option 1: If one (at end of exon 4/start of intron 4) is used, protein contains no hydrophobic stop transfer sequence, and protein is secreted.
b. Option 2: If other one (at end of exon 6) is used, protein contains hydrophobic sequence encoded by exons 5 & 6, and protein stays in plasma membrane.
3. mRNA can be spliced and/or poly A added in two alternate ways. Location of protein (antibody) depends on whether splicing of intron 4 or poly A addition happens first. Think of it as a competition. Either
a. Poly A adding enzymes get there before the spliceosome. In that case, poly A is added to site near end of exon 4, and rest of intron 4 (and rest of gene) is never transcribed, or
b. The spliceosome gets there first. In that case, Intron 4 is transcribed and spliced out before poly A can be added. (In this case, poly A is added at the end of exon 6 instead.)
4. Why are 2 forms of antibody needed?
a. Membrane-bound form of antibody: Serves as receptor for antigen = trap to detect when antigen is present. Binding of antigen (ligand) to antibody (receptor) serves as trigger to start secreting antibody.
b. Secreted (soluble) form: Acts as effector -- carries out major function of immune system -- binds to soluble antigen in body fluids and triggers destruction of antigen in multiple ways.
C. The general Principle -- You can get many different proteins from a single gene by the processes listed below. Therefore biologists are interested in proteomics (study of all proteins made in a cell) not only genomics (study of entire DNA or gene content).
1. Starting transcription at different points
2. Ending transcription (adding poly A) at different points
3. Splicing out different sections (exons as well as introns) of the primary transcript -- alternative splicing.
To review regulation & alternative splicing, try problems 4-13 & 4-14, & 4R-6.
IV. Regulation at translation.
A. How to control rate of translation? In principle:
1. Can regulate half life of mRNA (control rate of degradation).
a. In prokaryotes most mRNA's have a short 1/2 life; in eukaryotes this is not necessarily so.
b. Different eukaryotic mRNA's have very different half lives.
c. Note: proteasomes degrade only proteins NOT RNA's.
2. Can regulate rate of initiation of translation (control how effectively translation starts).
B. Some Famous Examples of Regulation of Translation. (The principles are important; we will not go into the details.)
1. Use of a regulatory protein: -- Have a protein that binds to mRNA (or some other part of the translation apparatus) and affects either initiation and/or degradation, depending on where it binds. Two examples:
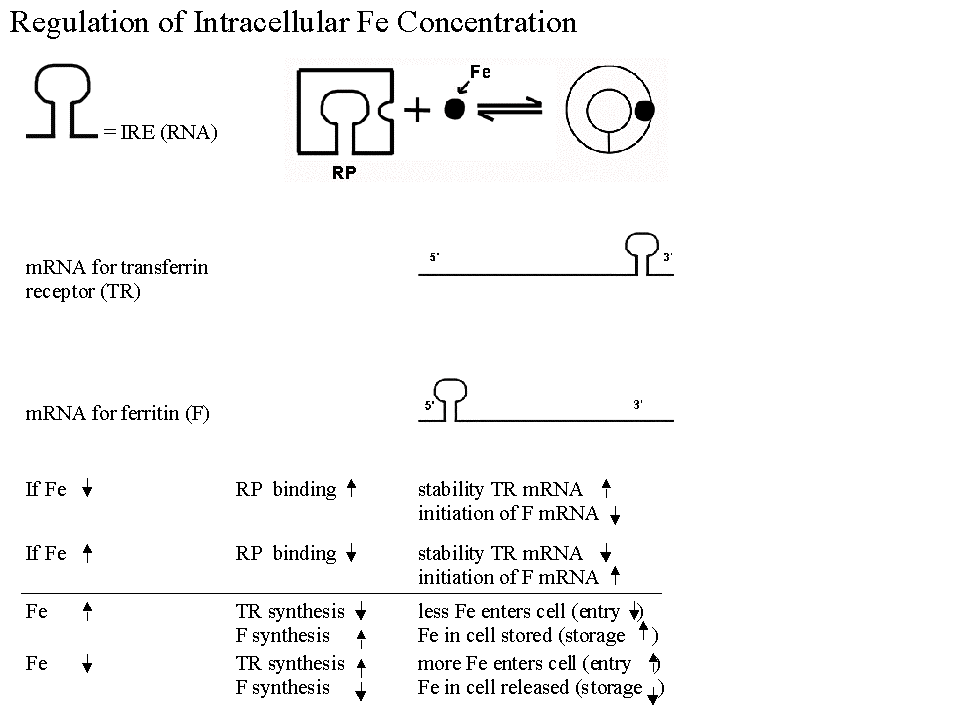
a. Regulation of synthesis of Ferritin & Transferrin Receptor (& intracellular iron levels).
- Regulatory protein acts like a (prokaryotic) repressor, but binds to regulatory sequence in mRNA, not DNA.
- Regulatory protein is allosteric, and level of small molecule effector (Fe) inactivates the regulatory protein.
- The regulatory protein binds to mRNA in the absence of Fe, not when Fe is high.
- Active form of repressor protein binds to more than one mRNA. Binds to mRNA for at protein A at 5' end (blocking initiation) and to mRNA for protein B at 3' end (blocking degradation).
- This is another example of coordinate control. There is one trans-acting factor here (repressor), but both mRNA's have the same cis acting sequence.
- See Becker, figs. 23-33 & 23-34 if you are curious about the details.
A question to think about: Given the information above, which is protein A, and which is protein B? Which one is ferritin and which one is the transferrin receptor? Ferritin is an intracellular protein that stores excess iron. You can check your answer in Becker or using this diagram.
b. Regulation of globin synthesis by heme (Becker fig. 23-32). Heme (the prosthetic group of hemoglobin) stimulates synthesis of globin (the protein part of hemoglobin). In this case, heme prevents inhibition of translation.
- In the absence of heme, inhibition occurs, and translation is blocked. No globin produced.
- Heme blocks the inhibition. Therefore, in the presence of heme translation proceeds. Heme relieves the block in translation, and globin is made.
Interesting features of this case worth noting are:
(1). Inhibition of inhibition results in stimulation; in other words, (-) X (-) = (+).
(2). Another example of coordinate control. This system ensures coordination between the supply of heme and of globin. Globin is useless without heme.
(3). FYI: Heme works by interfering with phosphorylation -- it blocks a kinase from phosphorylating a critical translation factor (ElF2 = elongation factor 2). Phosphorylation of ElF2 (in the absence of heme) inhibits ElF2 and blocks translation. This is another example of a protein (ElF2) that has active and inactive forms, and phosphorylation (or dephosphorylation) interconverts the two forms. (See V. A below.)
→ Formation of double stranded RNA. This triggers degradation &/or inhibition of translation of the mRNA.2. Use of a regulatory RNA -- RNA interference (RNAi)
a. Trans acting factors can be RNA. Not all regulatory factors are protein -- some are short RNA's. (These are usually derived from double stranded RNA -- See Becker figs. 23-35 & 23-36.)
b. How does a short RNA affect translation?
(1). Inhibition (usual case): Small RNA binds to mRNA
(2). Stimulation: Some recent cases have been discovered in which small RNA binds to mRNA and 'up regulates' translation. Mechanism so far unknown.
c. Use in Regulation: Cells naturally produce micro-RNA's that bind to mRNA's and regulate translation as above. The use of short regulatory RNA's to block translation appears to be important during regulation of development. (See Becker 23-36.)
d. Use in the Lab as a tool: Called RNAi = RNA interference. The use
of artificially added short double stranded (ds) RNA to block
transcription/translation and turn genes off is very common. (See Becker
23-35.) Enzymes of cell convert added ds RNA into short single stranded
RNA that interferes with translation and/or transcription as in b. Same
effect as adding antisense RNA (but works better).
V. Post Translational Regulation. Don't forget: regulation occurs after translation too -- after proteins are made, their activity can be modulated. Many examples of post translational modification have already come up and more will be discussed later. Here is a summary (mostly review):
A. Covalent Modification. Proteins can be modified covalently either reversibly (for ex. by phosphorylation and dephosphorylation), or permanently (for ex. by removal of N-terminal met., addition of sugars -- glycosylation, etc.)
See problem 6-3.
B. Noncovalent Modification. Proteins can be activated or inhibited by reversible noncovalent binding of other factors -- small molecule allosteric effectors, other proteins such as calmodulin (an important Ca++ binding protein to be discussed later), etc.
C. Degradation. Proteins can be selectively destroyed.
1. Half Lives Vary. Not all proteins have the same half life.
2. Proteasome: Major factor in regulation of protein turnover is control of addition of ubiquitin leading to destruction by proteasome. See Becker 23-38 or Sadava 14.24 (14.22) or the Nobel Prizes for 2004.
3. Significance: Important example of a family of proteins that all have a short half life = cyclins; control progression through cell cycle. Different cyclins control transitions from G1 to S, G2 to M etc. Cyclins are made as needed and degraded immediately after use. (Note: Both mRNA's for cyclins and cyclins themselves are degraded after use. More on this when we cover the details of the cell cycle.)
D. Location. Proteins can activated or inhibited by a change of location. For example, transporters like GLUT4 only work if positioned in the plasma membrane; if they are sequestered in vesicles they are inactive. Transport of glucose into the cell can be regulated by moving the GLUT4 in and out of the membrane.
To review post-transcriptional &/or post-translational regulation, try problem 4-15. By now you should be able to do all the problems in 4 & 4R except for a few fine points about hormones, which should be covered next time.
Next Time: How does
one cell signal to another?
{kind=link}