C2006/F2402 '09 -- Outline for Lecture 24
(c) 2008 Deborah Mowshowitz . Last updated 04/29/2009 10:02 AM.
Handouts: 24A = Top: Interactions of B & T cells (not on web); Bottom:
Stages of Immune Response (Bottom 1/2 = bottom 1/2 of class handout)
24B -- Structure
of Antibodies and their respective genes (posted version is labeled 24D)
Two interesting developments in genetics & physiology:
Some Athletes’ Genes Help Outwit Doping Test (5/30/08) and
I. Parts of the Adaptive Immune System
A. Specific Immune system has 2 branches
1. In both branches: Cells make a specific protein that binds to a foreign substance = antigen. Protein and antigen match up like ligand and receptor (or enzyme and substrate). Binding of specific protein to its target antigen is specific, and usually leads to destruction of target.
2. Humoral response -- Specific cell protein is an antibody. Why 'humoral?' Binding and destruction of antigen done by proteins in "humors" = antibodies in blood and secretions (for ex. milk, tears).
Example: B cells → release antibody → Ab (antibody) binds Ag (antigen -- usually on surface of microbe) → trigger destruction of microbes (microbes are engulfed by phagocytes or lysed) often with the help of a set of proteins called complement. (See Sadava 18.10 (18.11) & table below.) Allergies are a side effect of this system.
3. Cellular or cell-mediated response -- Specific cell protein is on surface of T cells, not released. Protein is called a TCR (T cell receptor). Binding and destruction of antigen done by whole cells bearing a TCR.
Example: T cells → TCR on surface; TCR's (of cytotoxic T cells) bind to Ag on surface of virus infected eukaryotic cell → destroy target cell by triggering apoptosis (programmed cell death). This is probably why grafts fail; foreign cells of graft look like infected (defective?) cells and are destroyed.
4. Big difference between the two branches -- Location of Target (as well as specific protein)
a. Humoral Response. Antibody (B cell protein) binds to antigens in solution or on surfaces of bacteria & viruses. Neither the protein mediating the immune response (antibody) nor its normal target (antigen) are on eukaryotic cell surfaces.
b. Cell-Mediated Response. TCR binds only to antigens on surfaces of other eukaryotic cells. Both the protein mediating the immune response (TCR) and its target must be on eukaryotic cell surfaces.
B. What Cells are involved? What are B cells and T cells? See handout 23C. White blood cells (leukocytes) -- contain no hemoglobin. WBC divided into two main types
1. Phagocytes -- macrophages, dendritic cells, etc. ( See Sadava fig. 18.2). Involved in processing antigens so lymphocytes can respond to them, and/or engulfing (& destroying) antigens identified by the immune system.
2. Lymphocytes. Found in lymph nodes and elsewhere. Do actual production of antibodies and/or execution of cellular immune response.
a. Divided into B and T cells.
(1). Both B & T cells come from same line of stem cells in bone marrow.
(2). B cells mature in bone marrow; T cells in thymus
b. Role of B cells -- produce & secrete antibodies. Major players in humoral response.
c. Role of T cells -- Needed for cell-mediated responses. Two types
(1). Helper T's (TH) -- Required for function of both TC's and B's .
(2). Cytotoxic T's (CTL or TC ) -- Kill target cells.
C. Table: Summary of Major features of 2 branches of specific immune system. Any features not covered yet will be covered later; this is here for reference. See notes after the table.
| Immune Response Type | Humoral | Cell-Mediated |
|
Cell involved in Response |
B cells |
T cells |
|
Protein Made by Cell |
Antibody (Ab) |
T cell receptor (TCR)* |
|
Location of Protein |
In serum, tears, etc. (released by B cell) or on B cell surface. |
Always on T cell surface (attached to T cell) |
|
Protein Recognizes |
Free Antigens (Ag) or Ag attached to microbial surfaces |
Antigens attached to surfaces of eukaryotic cells # |
|
Aide in killing targets (FYI) |
Complement** |
perforins## |
|
Usual targets (for killing) |
Microbes, soluble proteins |
Infected or cancerous cells (for Tc or CTL) |
|
Side Affect |
Allergies |
Graft rejection |
Notes:
*T cell receptor is NOT the receptor for T cells -- it is the
protein on the T cells that is the receptor for an antigen. It is the receptor of
T cells, not the receptor for T cells.
** Complement = a series of proteins found in blood.
Activation of complement involves a cascade of activations similar to that
involved in blood clotting. Complement binds to antibody-antigen complexes
attached to microbes and triggers phagocytosis or lysis of the microbe bearing
the complex.
# Antigen must be attached to a
euk. cell surface protein called MHC. (Details later.)
##Cytotoxic T cells use proteins called perforins to make holes in their targets. Then other proteins enter the holes and trigger apoptosis. Note complement is similar to perforins but works on prokaryotic invaders; perforins work on rogue eukaryotic cells. Many texts say perforin lyses cells -- it makes holes in membrane, and then water enters, causing cells to swell and burst. (This is the way complement kills bacteria.) Newer data indicates perforin works to trigger apoptosis.
II. How does Clonal
Selection Work? See Sadava fig. 18.6 (18.7)
A. Features that are same with both B & T cells
1. DNA rearrangement occurs during development -- therefore each cell has a unique set of genes for antigen binding protein.
2. DNA rearrangement occurs before exposure to Ag.
3. One protein with unique binding site made per cell. (Either one antibody/BCR or one TCR.) Note this means only one of two alleles of gene are expressed in this case.
4. Protein (BCR or TCR) has a variable part (specific for antigen) and a constant part (not dependent on Ag).
5. Mechanism of clonal selection -- Response to Antigen
a. How Ag 'selects' cells making the 'right' Ab or TCR -- Ag binding protein acts as trap/receptor for Ag. Antigenic determinant (epitope) binding to surface Ab (BCR) or TCR serves to select cells of appropriate specificity.
b. What accounts for tolerance and/or memory ? Clonal expansion (or suppression) occurs in response to Ag binding.
(1). Destruction. If Ag is perceived as "self" → cell (T or B) destroyed or suppressed (→ tolerance). See Sadava fig. 18.7 (8th ed. only)
(2). Activation. If Ag is perceived as foreign → cell divides → clonal expansion, further differentiation into effector or memory cells. (See below for details.)
(3). Whether antigen is perceived as "self" or "foreign" depends on time of exposure to the antigen (embryonic vs adult) and additional factors. (This turns out to be very complicated, so we are ignoring the "additional factors.")
B. Features that are unique to T cells (See top of handout 24A for B vs T, and TC vs TH )
1. Protein made by T cell is T cell receptor, not Ab. (See Sadava fig. 18.12 (18.13)). Each T cell makes a unique TCR (also called T cell antigen receptor) due to DNA rearrangements of TCR genes.
2. T cell receptor always remains on cell surface; never secreted
3. Antigen must be on eukaryotic cell surface:
a. Antibody will bind to free antigen in solution (or to part of a whole bacterium). TCR will not.
b. TCR only binds to Ag on surface of another (euk.) cell. .
c. TC vs TH
(1). TC
(a). Target cell: TC binds to ordinary (euk.) cell with abnormal epitopes on surface for example, an infected cell.
(b). Result of binding to target cell: TC destroys target.
(c). Surface marker: TC is CD8+ -- has protein called CD8 on surface
(2). TH
(a). Target cells: TH binds to immune system cell with abnormal epitopes on surface, for example, a B cell that has bound Ag.
(b). Result of binding to target cell: Binding activates the TH cell &/or B cell. (Promotes the immune response -- details vary depending on type of immune cell.)
(c). Surface marker: TH is CD4+ -- has protein called CD4 on surface
III.
Activation of B and T cells -- what triggers clonal expansion?
See Sadava fig. 18.15 (18.17) for overall picture.
A. Why is activation needed?
1. For Clonal Expansion: a B or T cell must be activated in order to divide and specialize, and to form both memory cells and effector cells. See Sadava figs. 18.6 & 18.15 (18.7 & 18.17).
2. What do effector cells do?
a. Effector B cells (plasma cells) secrete antibody
b. Effector TC cells kill targets
c. Effector TH cells provide juxtacrine and paracrine signals that promote the activation and functioning of other immune cells.
B. What is required? To activate a B or T cell, cell must get juxtacrine signals and paracrine signals. See Sadava fig. 18.14 (18.16)
1. Paracrine = a cytokine = secreted protein that affects development of the immune system & some related functions.
a. Terminology: Cytokines made by leucocytes often called interleukins, abbreviated IL-1, IL-2, etc.
b. Example of action: Activation of both B and TC cells requires paracrines from TH cells. See texts if you are interested in names and functions of various cytokines. (No details of paracrines will be covered in class; some details are included here and in problem book FYI only.)
2. Juxtacrine -- Involves contact between surface proteins on two cells -- a T cell & its partner (target cell). See handout 24A and Sadava fig. 18.14 (18.16). At least two juxtacrine interactions are required:
a. T cell must have: TCR & CD4 or CD8: CD4 (if it's a TH) or CD8 (if it's a TC)
b. Partner must have epitope plus MHC on its surface. (MHC = cell surface protein; details below).
(1). TCR binds to epitope -- provides specificity of Ag/Ab or Ag/TCR match
(2). CD4 or CD8 binds to MHC -- type of MHC distinguishes targets to be 'helped' (by TH) from those to be destroyed (by TC ).
c. Other proteins are involved too; we are ignoring them. Consult advanced texts if you are interested.
C. Role of MHC See Sadava fig. 18.15 (18.17)
1. What is it? MHC = very variable surface protein. (MHC is an acronym for major histocompatibility complex.) Related in structure to antibodies and TCRs. (For pictures of MHC molecules see picture from Alberts two types of MHC)
2. What does MHC do? MHC and small pieces of antigen (epitopes or antigenic determinants) form a complex. Complex is on cell surface, so epitopes are 'displayed' on the cell surface, stuck to the MHC molecules.
3. Two types of MHC
a. MHC I. All nucleated cells have MHC I on their surface.
b. MHC II. On cells of immune system only.
(1). Only certain cells of immune system (phagocytes & B cells) have MHC II on their surface.
(2). Not all T cells have MHC II at all times, & we will assume T cells do not have MHC II.
c. What proteins pair up?
(1). CD4 binds to MHC II
(2). CD8 binds to MHC I
d. What cells pair up? Two types of T's bind to different MHC's (w/ Ag) -- this is how T cells tell immune cells (that have captured Ag) and infected (ordinary) cells apart.
(1). Cytotoxic T's (CD8+) bind to target cells with Ag + MHC I on surface.
(a). TC are said to be "MHC I restricted" -- note target must have MHC I and Ag.
(b). Target cells for cytoxic T's are usually ordinary cells making abnormal proteins -- infected cells, for example.
(c). Binding to target (abnormal) cell → final activation of TC and killing of target cell. (Additional earlier steps of activation are required; we are ignoring them.)
(2). Helper T's (CD4+) bind to target cells with Ag + MHC II on surface.
(a). TH are said to be "MHC II restricted" -- note target must have MHC II and Ag.
(b). Target cells for helper T's are usually cells of the immune system, especially B cells and phagocytic cells that have internalized foreign antigens. These are called 'antigen presenting cells' or APC's.
(c). Binding to target cell (APC) → activation of TH.
(d). Activated TH can then activate a B cell or TC. In some cases, if APC is a B cell, activation can be mutual. (See 'one step' below.)
e. Activation of B cells (See Sadava fig. 18.15 (18.17))
(1). Two step:
(a). Phagocytic APC (not a B cell) binds to and activates a TH (See Sadava fig. 18.13 (18.15)).
(b). Activated TH detaches from APC; binds to and activates a B cell.
(2). One step: B and TH bind to and activate each other. B acts as APC to activate TH; TH in turn activates B.
D. More on MHC
1. Details (FYI)
a. Types: There are 2 main types, MHC I & MHC II, and many versions of each type.
b. Genes: Each individual has several different genes for each of the two main types of MHC. Each of these genes has 20-40 or even more variants (alleles).
c. Person to person variation: Since there are several genes per person and many different alleles of each gene in the population, there is a lot of variation in the actual MHC proteins (and DNA) from person to person.
d. No cell to cell variation: These genes, unlike genes for antibodies and TCR's, do not rearrange during development. All cells in person have the same MHC DNA. So there is variation from person to person, but all cells in a single person have the same MHC genes and make the same MHC proteins (multiple types made per cell).
2. How does epitope get on MHC? How are antigens 'presented' or 'displayed' on the cell surface?
a. How the pieces get to attached to MHC -- depends on type of cell and where protein comes from.
(1). Infected cells -- proteins made inside the cell are digested in proteosomes; protein fragments (epitopes) enter the ER using a special transporter, and bind to MHC I.
(2). Cells of immune system -- proteins made outside the cell are engulfed (by phagocytic cells) or endocytosed (after binding to antibody on surface of B cells). Protein fragments bind MHC II in endosomes.
b. How MHC + epitope reaches cell surface -- MHC's are transmembrane proteins in subcellular membranes (of endomembrane system); MHC's bind epitope and complex reaches cell surface by exocytosis.
c. Many epitopes displayed per cell -- every APC (antigen presenting cell) 'presents' many different pieces of whatever antigen(s) it engulfed, endocytosed or made.
Try Problems 13-5 & 13-9. For a
review of the information so far, try 13-6 , 13-11 (skip C) & 13-12.
IV. How do
T cells get activated? Summary Table. See also Sadava, fig. 18.14
(18.16).
|
What's Activated? |
Antigen Presenting/Target Cell |
What holds Epitope |
Source of Antigen |
Result |
| Cytotoxic T | Infected Cell |
MHCI |
Made in infect. cell |
Killing of Target Cell; |
| Helper T | Classic APC (B, macrophage, etc.). |
MHCII |
From outside APC |
Mitosis of TH cell to give clone |
Notes:
(1). Activation of lymphocytes also requires appropriate
cytokines. TH cells need IL-1 from the APC's; TC cells
need IL-2 from TH and B cells need various IL's; class of Ab made by
B cell depends on type of IL it gets.
(2) In a TH -- B cell combo, each can activate the other. Alternatively, a helper T can be activated first, and then activate a B cell.
V. Ab
Structure -- See handout 24B, picture of an
immunoglobulin (from Alberts), or Sadava fig. 18.9 (18.10). What is the
molecular structure of antibody molecules?
A. V vs C -- types of Immunoglobulin (Ig).
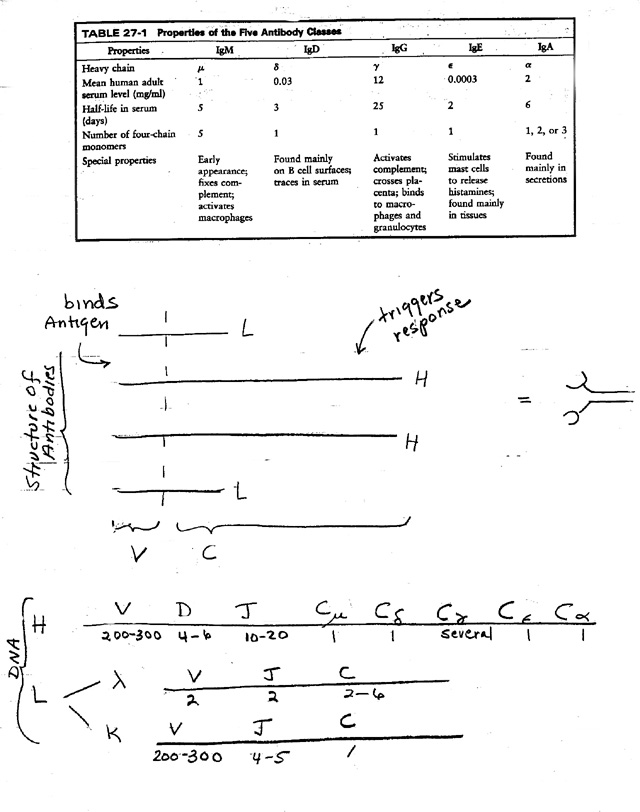
1. There are 5 main classes of Antibody -- IgM, IgD, IgG, IgE, and IgA. See table on handout 24B & Sadava Table 18.3.
2. V & C: Each Ab or Ig is made up of a V section ("variable" region or Vee) & a C section ("constant" region or Cee).
3. Variable region
a. V is specific for Ag (or epitope). Determines what Ag will be bound = grabbers.
b. V is variable due to differences in sequence, not just differences in folding around Ag.
c. Every Ab or Immunogloblin (Ig) has (at least) 2 grabbers.
d. All grabbers in one Ab are the same.
e. All the antibodies made by one Ab-producing cell have the same V. All the antibodies made by descendents of that cell have very similar V's. (Minor differences are due to somatic mutation; see advanced texts if you are interested. We will ignore somatic mutation for the rest of this discussion, and assume all the antibodies made by the descendents of one cell have the same variable region.)
4. Constant region
a. C determines biological effects -- localization of Ab, and what will happen as consequence of binding Ag. (Whether complement will be activated, whether Ab will be found primarily in blood or secretions, etc.)
b. 5 main types of C regions, therefore 5 main classes of antibody. (For properties of the dif. classes see handout 24B or Sadava Table 18.3 )
c. The same V's can go with different C's. (Called "Class Switching")
(1). All the antibodies made by one Ab-producing cell do not necessarily have the same C.
(2).The antibodies made by descendents of a single cell may have different C's. The same variable region can go with different constant regions as B cell clone expands. How is this possible? Need a closer look at Ig structure
B. H vs L. See handout 24B or Sadava fig. 18.9 (18.10)
1. Every Ig has 2 kinds of chains, L ("light") and H ("heavy"). Light and heavy refer to relative differences in mol. wt.
2. Basic unit is 2 of each for a total of 4 chains. (For number of basic four-chain units per Ig, see table.)
3. Variable region (grabber) made of parts of each.
4. Each chain has a constant region
a. 2 kinds for L (kappa or lambda)
b. 5 basic kinds for H (mu, delta, gamma, epsilon or alpha)
c. Hc (constant part of H) determines class (IgM, IgD, IgG, IgE, IgA)
d. Class (determined by Hc) determines location & other aspects of function (see "special properties" in table)
d. Class switching involves the H chains only, not the L chains.
5. Myelomas & Hybridomas: Ig structure was figured out by studying proteins made by myeloma cells (cancers derived from Ab-producing cells) or hybridomas (hybrids of Ab-producing normal cells and cancer cells). Only way to get large numbers of cells all making the same Ab/Ig. See texts for significance of hybridomas and monoclonal antibodies. (Sadava 18.11 (18.12))
C. Classes of Ab and class switching during development of immune response
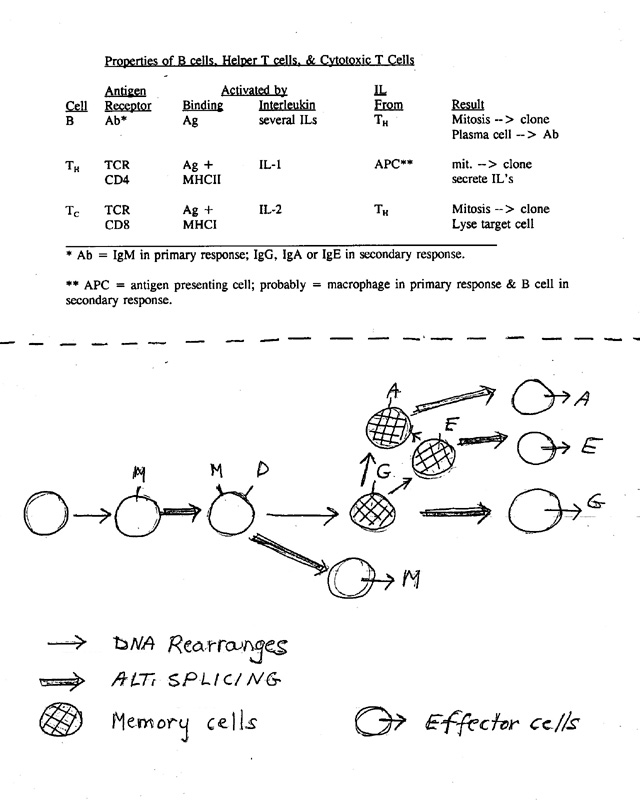
1. Order of events (see handout 24A, bottom) during immune response as B cell matures
a. First make M, then M + D -- all on surface.
b. Meet Ag → primary response: secrete M.
c. Meet Ag a second time → secondary response; secretes usually G but can be E or A.
d. All these Ig's combine with same Ag -- only constant region of H chain switches.
2. Implications of structure and switching
a. Can make different variable regions -- zillions of them, one for each dif. epitope. So the IgG, for example, in a person is a mixture -- all IgG molecules have the same constant regions but have different variable regions.
b. During different stages of immune response, can make Ab with same variable region but different constant region (for H). Can switch class and/or secreted vs. surface. How is this possible?
3. What we already know: How switch from membrane bound to secreted works. Variable part stays same; Hc changes from hydrophobic to hydrophilic by alt. splicing/poly A addition.
4. What we don't know so far: How do you make so many dif. variable regions AND What changes when you switch classes (from IgM to IgG? M to M + D)? Must be rearrangement of DNA or alternate splicing of RNA. Different solutions at different steps.
Try Problems 13-1 to 13-3.
VI. Structure of the DNA coding for Antibodies -- Basis of Generation of Diversity (G.O.D) and Class Switching
A. Basic idea: genes for H and L are mosaic -- Each "Gene" has several parts. See texts or handout 24B or Sadava 18.16 (18.18.)
B. How "gene" is divided -- region coding for each chain (H or L) has parts coding for each type of constant region and several parts coding for the variable region .
C. How DNA is used to make different antibodies (With different V's) -- DNA is rearranged
1. Pre Ag
a. Rearrange V/D/J region of DNA to make one coding region for variable part of H chain per naive/virgin B. Put one V next to one D next to one J. Delete all the intervening segments.
b. A similar process of DNA rearrangement occurs in DNA coding for variable part of L chain.
c. Net: Only one H chain allele and one L chain allele are rearranged and used. Therefore each cell makes only one type of variable region.
2. FYI: Post Ag -- Somatic Mutation** → minor changes in region of DNA coding for V regions of H & L chains. (No change in DNA coding for C regions ). In the secondary response, there is a second round of clonal selection for B cell variants making 'better Ab' -- Ab that binds Ag better (higher affinity Ab). This is why Ab made in secondary response is better at binding Ag than primary Ab.
3. Reminder: Switching at DNA level is unique to immune system. Parts of genes for antibodies (or TCRs) can be rearranged using enzymes that cut DNA and join (recombine) DNA segments that were not contiguous in the germline DNA. These enzymes are restricted to certain immune cells.
**Note: We are going to ignore the effects of somatic mutation, but it is included here for reference.
D. Summary of G.O.D (generator of diversity) -- how get so many V's?
1. H & L mix and match -- any H chain can go with any L chain
2. Mosaic V genes -- V parts (V, D, J) of DNA coding for each chain mix and match
3. Joins are inexact -- bases can be added when you rearrange the DNA -- when join V to D etc.
4. Somatic mutation -- post Ag
E. TCR genes are similar, except no somatic mutation. Genes are mosaic, and are rearranged. The proteins that are made have more than one chain; each TCR has constant and variable regions. See TCR picture from Alberts.
F. Class Switching -- How DNA is used to make different versions of the same antibody (with different C regions)
1. Definition of Class switching -- cell makes antibody with same variable region and different constant region.
2. Mechanism -- Switching occurs at DNA and RNA levels.
3. Pre Ag -- M vs D
Alternate splicing allows cell to produce M and D antibodies with same V/D/J (but different constant region of H chain). For details see Sadava fig. 18.17 (18.19).
3. Post Ag -- Alt splice of RNA and/or further rearrangement of DNA → new mRNA → new version of antibody with same variable region. Can have either of the following:
a. Rearrangement (usually deletion) of DNA → gene for H chain with original variable region and a new "constant" region. Make new class of antibody. (See Sadava fig. 18.18 (18.20))
b. Alternative splicing → mRNA for secreted version of cell surface antibody -- Same H chain except it's missing part that anchors the protein in the plasma membrane. Go from making "BCR" to making secreted antibody.
Try recitation problem 13-3 & problem 13-13 (Ignore IL-2 receptors).
VII. Evolutionary Aspects (FYI)
A. Clonal vs. Natural Selection. Note how clonal selection and natural selection compare. In both cases, need to have many variants (diff. antibodies or dif. organisms) to be able to respond to unpredictable environmental challenges. How is this done? In both cases, make many variants and conditions select (promote propagation of) cells making the few suitable Ab (or carrying out a rare, useful function); the rest are wasted. Random generation of variants seems wasteful, but is the biological solution to preparing for change without conscious planning ahead.
B. The Major Proteins of the Immune System are Related
1. The immune system uses 3 types of proteins that have a common evolutionary origin. These are antibodies, TCR and MHC. For additional pictures see Sadava fig. 18.9 (18.10) for antibodies & Sadava fig. 18.12 (18.13) for TCR. Here are links to parallel pictures (from Alberts) of the two types of MHC, a TCR, and an immunoglobulin (showing the domains).
2. All 3 types of proteins have a "constant" part and a "variable part."
a. Constant part determines where protein is (cell surface? What kind of cell? etc.) and its general function.
b. All 3 proteins bind epitopes -- Variable part determines what antigen/epitope will bind to the protein.
3. All 3 proteins include one or more copies of the immunoglobulin domain -- a section of the protein that is similar in structure and function. this is a common theme -- the same domains are found over and over in different proteins. (Examples are SH2 domains; DNA binding domains, etc.)
4. Variable part of antibodies and TCR's are generated by rearranging the DNA; the variable part of MHC's is encoded in the germ line -- the DNA inherited in the zygote is the DNA used to code for the MHC's. The DNA from MHC is NOT rearranged. However the genes for MHC's are polymorphic (have many different common alleles).
VIII.
Summary of Major Players in the Immune System:
| Cells | B cells, TC cells, TH cells, phagocytic cells, APC's |
| Secreted Proteins | Antibodies (Ab or immunoglobulins; 5 classes), Perforin*, Cytokines* |
| Cell Surface Proteins | MHC, BCR, TCR, CD4, CD8 |
The chart above summarizes the major players in immunology. You should be able to describe what each item is, its significance, and how it is related to all the others. "Secreted proteins" refers to those made by B and T cells. Proteins involved in the immune response (such as complement*) that are not made by lymphocytes are not listed. See ans. to problem 13-6, & tables above.
*Terms with a star have not been discussed in detail, but you should be aware of their general roles. You will not be asked any questions on the final about perforin, complement, somatic mutation, or which cytokine (or IL) does what.
Next Lecture (Optional) will cover regulation of the cell cycle, growth, and cancer.
{kind=link}
{kind=link}