C2006/F2402 '11 OUTLINE OF LECTURE #15
(c) 2011
Dr. Deborah Mowshowitz, Columbia University, New York, NY.
Last update
03/24/2011 02:16 PM.
Handouts: 14A --
Overview of Signaling -- the
Biological Big Bang Theory
14B --
Structure of G proteins and GPCR's;
cAMP pathway
15A --
Lining of the GI Tract & Typical Circuit
15B --
Homeostasis)-- Seesaw view for Glucose and Temperature Regulation;
I. How do Intracellular Receptors Work? (cont). See Sadava fig. 7.9 (15.8)
A-E. See last lecture.
F. Example -- Estrogen (A steroid)
1. Basic Mechanism. E → binds to estrogen receptors → complex; complex binds to estrogen response elements (EREs) in regulatory regions of (multiple) target genes. Binding → increased transcription of some genes (genes activated); decreased transcription of others (genes repressed).
2. Example of some proteins controlled by E -- controls production of receptors for other hormones. For example, during pregnancy controls production of receptors for oxytocin (in uterus) and prolactin (in breast). Oxytocin controls birth contractions; prolactin controls milk production.
a. In uterus: estrogen binding → activates transcription of gene for oxytocin receptors → production of new receptors for oxytocin = up regulation of oxytocin receptors. Receptors needed to allow response to contraction signal (oxytocin) → contractions → birth.
b. In breast: estrogen binding → inhibits transcription of gene for prolactin receptors → down regulation of prolactin receptors; at birth, estrogen level falls and inhibition stops → transcription of gene for prolactin receptors → synthesis of prolactin receptors → response to lactation signal (prolactin).
|
Summary of Effects of Estrogen |
||
| Target Organ | Uterus | Breast |
| Affects Receptors for | oxytocin ( → contractions) | prolactin ( → lactation) |
| Receptors are | up-regulated | down-regulated |
| Effect | Response to oxytocin increases | No response to prolactin |
| Result | Contractions & birth possible | Lactation only after birth (when estrogen levels fall) |
3. Why different results (different patterns of transcription) in different tissues -- in response to the same lipid soluble hormone?
a. May be different hormone receptors in different tissues. Many hormones/signals have multiple types of receptors. (Examples another time.) That's not the explanation here -- in this case, receptors in both cell types are the same.
b. Combinations of TF's (& factors that affect state of chromatin) in each cell are different; often more than one TF is required to get proper transcription of each gene. Hormone signal acts as trigger, but type of hormone effect (what is triggered) depends on other factors.
c. Reminder: All cells have the same DNA. Therefore
- All cells (except immune system) have the same cis acting regulatory sites -- the same HRE's, enhancers, etc.
- It's the trans acting factors such as hormone receptors that vary, not the cis acting regulatory sites.
- All cells have the same genes for the trans acting factors, receptors etc., but different genes are used to make regulatory proteins in different cells.
Other examples of how hormones can give different results in different tissues will be discussed later. In general, what any hormone does depends on combination of proteins (enzymes, TF's, etc.) already in target cell.
Try problem 6-19. By now you should be able to do 6-12 to 6-15.
II. Types of Cell Surface (Transmembrane) Receptors (See bottom of 14A)
A. Channels. Some receptors are themselves (parts of) channels. See AcCh receptor in previous lecture and Sadava fig. 7.6 (15.5) Other receptors are not channels, but work by opening or closing separate channels. Channels will be discussed next week by Dr. Firestein.
B. Types of Cell Surface receptors that are not channels
1. Type 1: Linked to G proteins.
a. Terminology: Called G protein linked receptors, or G Protein Coupled Receptors (GPCRs). For a generalized case, see Sadava fig. 7.8 (15.7).
b. Structure: All are 7 pass transmembrane proteins with same basic structure; all belong to same protein/gene family. (See Becker 14-4 or top of handout 14B.)
c. What are they receptors for? Many hormones such as TSH & epinephrine use GPCRs.
d. How do they work? Activate a G protein, which acts as a switch to trigger amplification (details below). G proteins usually:
(1). Activate enzymes that generate second messengers (see Sadava fig. 7.8 (15.7)), or
(2). Open/close ion channels.
2. Type 2: Not linked to G proteins. To be discussed in more detail later when we get to cell cycle and cancer. For reference:
a. Many are protein kinases. In addition to extracellular, ligand binding domain, have an intracellular kinase domain, or interact with an intracellular kinase (when activated).
b. Most well known type: Receptor Tyrosine Kinases (RTKs) -- also called TK linked receptors.
c. Structure: Usually are single pass proteins that aggregate into dimers when activated.
d. What are they receptors for? Many Growth Factors use TK linked receptors or related receptors. (See Becker table 14-3 if you are curious).
e. How do they work? These usually generate cascades of modifications, but do not always use 2nd messengers. If you want to see an example, see Sadava figs. 7.6 & 7.12 (15.6 & 15.10). We won't get to details of how these work for a while.
III.
G proteins -- How do they Fit In? How do they work?
A. What are the important properties of G proteins? (See Becker fig. 14-5 & Handout 14B)
1. Have active and inactive forms
a. Active form is bound to GTP
b. Inactive form is bound to GDP.
2. G-proteins act as switches in many processes (not just signaling)
a. Activation: G protein is activated by dumping GDP and picking up GTP in response to some signal.
b. Inactivation: G protein inactivates itself by catalyzing hydrolysis of GTP to GDP.
c. Why is it a switch? The G protein does not stay active for long. "Turns itself off."
B. Typical Pathway -- Role in signaling (see also handout 14B, middle panel)
| ligand (1st messenger) binds outside cell | → | activate receptor in membrane | → | activate G protein in the membrane | → | activate target enzyme in membrane | → | generate small molecule (2nd messenger) inside cell |
Note that the ligand (1st messenger) binds to the extracellular domain of its receptor. The remaining events are inside the cell. More on 2nd messengers below.
C. Activation & Inactivation of G Proteins
→ Protein-GTP (active) + GDP1. GTP exchange: Mechanism of activation & inactivation
a. Activation Reaction (GTP/GDP exchange, NOT phosphorylation of GDP; GTP replaces GDP):
Protein-GDP (inactive) + GTP
b. Inactivation Reaction (hydrolysis of bound GTP to GDP):
Protein-GTP (active) → Protein-GDP (inactive) + phosphate.
c. Overall: GTP displaces GDP, activating the G protein; GTP is then hydrolyzed (usually rapidly), returning the G protein to its inactive state.
(1). Net effect on GTP -- GTP hydrolysis: GTP ( + water) → GDP + phosphate
(2). Net Effect on G protein -- none: Protein is temporarily activated, but then inactivated. However protein cycles from inactive to active and back to inactive -- acts as switch.
d. Terminology. Since the overall result is that GTP is hydrolyzed to GDP, G proteins are sometimes called "GTPases."
2. What triggers activation?
a. General Case: Binding of a protein called a GEF (guanine-nucleotide exchange factor) causes GDP to fall off, and GTP binds.
b. In signaling: Activated GPCR = GEF. Binding of activated receptor to G protein triggers activation of G protein (causes loss of GDP).
3. What triggers inactivation?
a. G protein itself has enzymatic activity -- catalyzes inactivation (hydrolysis).
b. No trigger required -- hydrolysis of GTP to GDP happens automatically.
c. Other proteins may increase speed of hydrolysis. They are called RGS proteins (Regulators of G protein Signaling) or GAP proteins (GTPase Activating Proteins).
D. Types of G proteins
1. Subunits -- Ordinary G proteins are trimeric = they have 3 subunits.
a. Inactive G prot = heterotrimer of alpha, beta, gamma
b. Separation occurs on activation: On activation, alpha subunit (with the GTP) separates from other 2 subunits.
c. Either part may be the effector that actually acts on target -- alpha, or beta + gamma, can act as activator or inhibitor of target protein.
d. Hydrolysis causes reassociation. Hydrolysis of GTP to GDP causes alpha to reassociate with other subunits → inactive heterotrimer
2. Small G proteins -- to be discussed further when we get to cell cycle & cancer. For reference:
a. Structure: Small G proteins have no subunits.
b. Example: the protein called ras -- important in growth control; many cancer cells have over-active ras.
c. Role of GTP/GDP exchange: Are activated by GTP/GDP exchange, and inactivated by hydrolysis of GTP to GDP, as above.
d. Are not activated by GPCRs directly (other 'middle man' adapter proteins are involved)
3. How many G proteins?
a. There are many different G proteins. G proteins are involved in a very large number of cellular processes, not just signaling. (We have ignored their importance until now. See Becker for details & many examples. )
b. Active G proteins can be inhibitory or stimulatory.
c. Method of action: Activated G proteins work by binding to and activating (or inhibiting) other target enzymes/proteins.
d. Terminology: The different trimeric G proteins are usually known as Gp, Gq Gi, Gs etc. (Books differ on details of naming.)
E. For reference: Comparison of Protein Kinases, Receptor Protein Kinases, & Trimeric G proteins
Protein Catalyzes What's added to Target Protein? Who gets the P or GTP? How Inactivated? Protein Kinase Protein + ATP → ADP + protein-P Phosphate Usually a dif. protein Separate Phosphatase removes P Receptor Protein Kinase** Protein + ATP → ADP + protein-P Phosphate Usually separate subunit of self Separate Phosphatase removes P Trimeric
G ProteinExchange & Hydrolysis as described above. GTP Itself Hydrolyzes GTP to GDP (by itself)
**Receptor protein kinases have an extracellular ligand binding domain and an intracellular kinase (or kinase binding) domain. Ordinary kinases usually add phosphates to other proteins. Receptor kinases usually add phosphates to themselves. (For an example, see Sadava fig. 7.7 (15.6)
Try problems 6-1 & 6-2.
III. 2nd Messengers -- How do they fit in? How do they work?
A. Typical Pathway -- where does 2nd messenger fit in? (see also handout 14B, middle panel)
| ligand (1st messenger) binds outside cell | → | activate receptor in membrane | → | activate G protein in the membrane | → | activate target enzyme in membrane | → | generate small molecule (2nd messenger) inside cell |
B. What are Second messengers? -- See handout 14B or Sadava fig. 7.8 (15.7)
1. What are they? Small molecules or ions that move through the cell and bind to their target proteins.
2. The usual second messengers -- see handout 14B for structure of cAMP and mode of action
| 2nd Messenger | Where does it come from? | How is it made? |
| cAMP | ATP | by action of adenyl cyclase |
| DAG & IP3 | membrane lipid | by action of phospholipase C |
| Ca2+ | stored Ca2+ in ER (or extracellular) | by opening channels (in ER/plasma memb.) |
3. What do 2nd messengers do? Bind to and thereby activate (or inactivate) target proteins.
4. How they are made: Active G protein (subunit) → binds to & activates enzyme in (or associated with) membrane → generates second messenger in cytoplasm. See Becker fig. 14-7 or Sadava figs. 7.8 & 7.14 (15.7 & 15.12) for cAMP pathway. We will get to IP3 pathway later. If you are curious, see Becker fig. 14-10 or Sadava fig. 7.15 (15.13).
5. A Specific Example: Thyroid stimulating hormone (TSH) -- promotes release of thyroid hormone (TH).
a. Generation of 2nd messenger (cAMP)
TSH (1st messenger) binds → activate GPCR in membrane → activate G protein in the membrane → activate enzyme in membrane (adenyl cyclase) → generate small molecule (2nd messenger) inside cell = cAMP b. Action of 2nd messenger
cAMP → activate protein kinase (PKA) in cytoplasm → phosphorylate target enzymes → stimulate multiple steps in synthesis and release of TH
6. Why bother with all these steps?
a. Amplification. Many steps involve amplifications. For example, one molecule of active Adenyl cyclase can generate many molecules of cAMP and one molecule of PKA can phosphorylate many molecules of its target enzymes. For an example of the possibilities of a cascade of amplification, see Becker fig. 14-3 or Sadava fig. 7.20 (15.18). (They don't agree on the exact numbers.)
b. Examples. The usual example for this type of modification cascade is the breakdown of glycogen, stimulated by the hormone epinephrine (adrenaline), which is the example in Becker fig. 14-3. This example was the first to be discovered, but is more complex than the TSH case.
If you are interested in the details or want to read ahead, see Sadava fig. 7.20 (15.18) or Becker figs. 14-25 (14-24) & 6-17 (6-18 ), or the handout at http://www.columbia.edu/cu/biology/courses/c2006/handouts/glycogen09.pdf.
IV. An example of
a second messenger -- cAMP & its target (PKA) See
handout 14B or Becker fig. 14-7.
A. How is cAMP level regulated? What does it do?
1. How is cAMP made?
a. G protein activates adenyl cyclase (also called adenylyl cyclase or AC)
b. cAMP made from ATP by adenyl cyclase; for structure of cAMP see handout and Becker fig. 14-6 or Sadava fig. 7.14 (15.12).
2. What does cAMP do? See Becker table 14-1 (14-5).
a. cAMP binds to and activates protein kinase A = PKA. (Also called cyclic AMP dependent protein kinase = cAPK) See Becker fig. 14-8.
b. PKA adds phosphates to other proteins
(1). Phosphorylation by PKA can activate or inhibit target protein (target = substrate of PKA)
(2). PKA action can modify other kinases/phosphatases and start a cascade
(3). End result varies. Depends on which kinases and phosphatases in that tissue are targets (substrates) of PKA and/or the other kinases/phosphatases (at end of cascade). See example below.
3. How does signal system turn off when hormone leaves?
a. G protein doesn't stay activated for long: Activated G protein hydrolyzes its own GTP → GDP (→ inactive G protein).
b. cAMP is short lived -- it's hydrolyzed by phosphodiesterase (PDE)
c. In absence of cAMP, action of kinases are stopped and/or reversed
(1) PKA becomes inactivated
(2) Phosphatases become active -- remove phosphates added by kinases
B. How do hormones work through cAMP?
1. TSH -- see above. PKA phosphorylates (and activates) enzymes needed to make thyroid hormone.
2. Glycogen metabolism: This case is very complex and will be discussed later. See above for references.
V.
Introduction to Physiology & Multicellular organisms -- What is signaling
good for?
A. Single cell Life Style vs. Multicellular
1. Single celled organisms
a. Surrounded by external environment -- Can't change or regulate it
b. Have one basic function -- grow and multiply
c. Respond to external conditions (since can't change them) to maintain optimal intracellular state
(1). Pick up and/or dump what is necessary for metabolism
(2). Keep intracellular conditions (pH, level of amino acids, oxygen, etc.) as constant as possible and expend minimal energy by adjusting rates of transcription, enzyme activity, etc.
d. Note no specialization: each cell does all possible functions
2. Multicellular organisms & Homeostasis
a. Each cell in organism surrounded by internal environment. (See Sadava fig. 40.1, 9th ed.) Extracellular fluid (ECF) that makes up internal environment is composed of:
plasma = liquid part of blood = fluid between blood cells
interstitial fluid (IF) = fluid between all other cells
b. Organism as whole can regulate composition of internal environment (milieu); therefore can maintain relatively constant external environment for each cell. Process of maintaining a relatively constant internal environment (of whole organism) = homeostasis.
c. Each cell has two basic functions
(1). Grow or maintain itself as above
(2). Specialized role in maintaining homeostasis of whole organism
d. Cells are Specialized. Maintenance of homeostasis requires co-operation of many different cell types, not just circuits within a single cell.
Summary of Above:
| Unicellular Organisms | Multicellular Organisms | |
| What surrounds cell? | External environment | Internal environment of organism |
| Can organism regulate what surrounds each cell? | No | Yes |
| How many functions of each cell? | 1 | 2 or more |
| Is cell specialized? | No | Yes |
B. Organization -- How are cells set up to co-operate in a multicellular organism? See 15A.
1. Cells, Tissues & the 4 major tissue types (5, if you count the blood separately) -- see lecture #4, & Sadava fig. 40.7
2. Organs
a. Made of (different kinds of) tissues.
b. Example: lining of GI tract. Has layers of different tissues -- epithelial, connective, muscle, and nervous; these serve primarily for absorption (of material from lumen), support, contraction, and regulation respectively. (See handout 15A or Sadava fig. 40.7) The blood (a type of connective) doesn't really fit in this classification -- serves for transport of materials in and out.
3. Systems -- Group of Organs → body or organ system. Work together to maintain homeostasis for some component. See Sadava 40.1 (8th ed). Number of systems depends on who's counting. Usual # is 8-12.
a. Immune system -- responds primarily to internal changes caused by presence of foreign organisms (or their macromolecules) -- responds to viruses, bacteria, cancer cells, etc. (Graft rejection, allergy, etc. are side effects of this.)
b. Other systems -- respond to changes in internal mileu caused by other factors.
VI. How
is a component of the internal milieu regulated?
A. General Principle -- Homeostasis is maintained by Negative Feedback
1. What is negative feedback? The system is self correcting -- it responds so as to decrease deviations from the set point. Deviations in either direction (too high or too low) are corrected back to standard (the set point).
2. How is negative feedback different from positive feedback? In positive feedback, the system responds so as to increase deviations from the set point -- a small deviation triggers a bigger one, which triggers a bigger one and so on. The deviations get bigger and bigger until → boom! (Contractions leading to birth, summary of graded potentials until you generate an AP, lactation, etc.)
3. Results of negative FB: Value of regulated variable (blood glucose, or temperature) does not remain exactly constant, but stays within narrow limits.
4. Note on Terminology Some of the terms discussed here are used differently in molecular biology and in physiology. Fortunately, the meaning is usually obvious from the context. For example, the terms "effector" and "negative feedback" are used differently in the two contexts.
In physiology, negative and positive feedback are defined as above. For negative fb, it doesn't matter whether the corrections are achieved by inhibition (turning off the heater; stopping glucose production) or acceleration (turning on the air conditioner, increasing glucose uptake from blood).
In biochemistry, negative feedback usually means inhibition of an earlier step (& positive fb usually means activation).
'Effector' is also used differently in physio and biochem; see below.
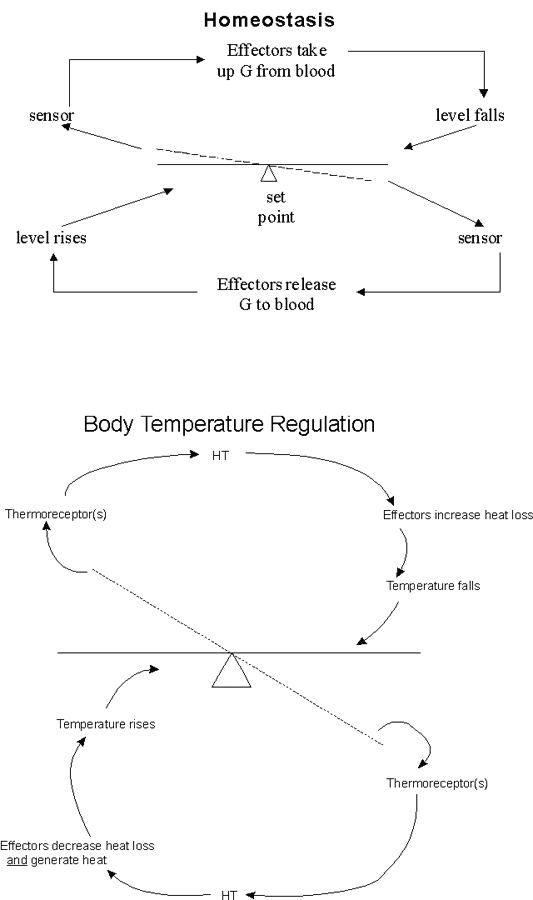
B. Example #1 -- Regulation of blood glucose levels. The see-saw view. See handout 15B or Sadava fig. 51.18 (50.19). The fig. in the 9th ed. is not a see-saw, but the point is the same.
1. Have a regulated variable -- glucose level in blood.
2. Need a sensor (or receptor) -- to measure levels of "regulated variable" (glucose). Here, sensor is in pancreas.
3. Need effector(s) -- to control levels of regulated variable (glucose) -- usually have one or more effectors that respond in opposing ways. In this case, effectors for uptake of glucose are liver, adipose tissue, and skeletal muscle; effector for release of glucose is liver.
Note on terminology: In physiology, "effector" usually means "a tissue or organ (like muscle or liver) that carries out an action and thus produces an effect." In this example, the effectors = organs that act to raise or lower the blood glucose. In molecular biology, the term "effector" is usually used to mean "a modulator of protein function." A modulator = a small molecule (like an inducer, enzyme activator etc.) that binds to a protein, alters the shape and/or function of the protein, and thus triggers an effect.
4. Have a set point -- the level the regulated variable (blood glucose) should be. Set point is also sometimes used to mean the level at which corrections (to raise or lower the value) kick in.
In most cases, there is no significant difference between these two definitions of set point. In some cases, the desired value (first definition) and the value at which corrections occur (second definition) may be different. For example, there may be two cut-off points-- upper and lower, that bracket the desired level of a regulated variable. At levels above or below the respective cut-off points, messages are sent to the appropriate effectors to take corrective action. The term "critical values" is sometimes used instead of "set points" to describe the cut-off point(s).
5. Signaling -- need some signal system to connect the sensor(s) and the effector(s). Can be nervous &/or hormonal. In this case, primary (but not only) signal is hormonal & primary hormones (signals) are insulin & glucagon.
6. Operation of Negative Feedback -- the system responds to negate deviations from the set point. Important features:
a. Works to stabilize levels of blood glucose (the regulated variable)
b. System is self-correcting -- Deviations in either direction (if blood glucose is either too high or too low) are corrected.
c. There are two opposing actions by effectors, not just one.
(1). If [G] gets too high, effectors take G up from blood. (top half of seesaw diagram)
(2). If blood [G] gets too low, effector releases G to blood. (bottom half of seesaw diagram)
d. Negative feedback is not always inhibition. The deviation from the set point may be fixed by accelerating, not inhibiting, a process. In negative feedback, deviations from the set point can be corrected either by speeding up a process (such as glucose uptake) or slowing down a process (such as glycogen breakdown to glucose). For example:
(1). If blood [G] goes up, uptake from blood increases and glycogen breakdown decreases. In this case, an increase in glucose uptake is used to help decrease the deviation from the set point.
(2). If blood [G] falls, release into blood increases, and glycogen synthesis decreases. In this case, an increase in glucose release is used to help decrease the deviation from the set point.
(3). In both cases, one process is increased and another is inhibited to help decrease the deviation from the set point.
7. Net Result -- regulated variable ([G] in blood) is not constant, but stays close to set point.
See problem 5-1 & 5-2 a & b.
C. Example #2 -- Regulation of body temperature (in humans) -- the see-saw view (handout 15B)
1. Note many features are same as in glucose case. (Can you list them??)
2. Features not found in glucose case:
a. Multiple sensors in different places (for core and skin temp.). How to integrate multiple inputs?
b. Nature of Signal -- Signals are neuronal, not hormonal
c. Integrative center (IC)
(1). Role of IC: Compares set-point to actual value, sends appropriate message to effectors.
(2). Type of IC
(a). Sensor/IC function may be combined, as in Glucose example.
(b). Separate IC needed if there are multiple sensors, as in this case. IC co-ordinates incoming information from multiple sensors
(3). In this example, IC = hypothalamus (HT)
3. Organs/body systems involved as effectors
|
Effector |
Action To Raise Temp |
Action To Lower Temp |
|
Skeletal muscles |
Contraction generates heat (shivering) |
None |
|
Smooth muscle of peripheral blood vessels in skin |
Muscles contract; vessels constrict to reduce heat loss |
Muscles relax; vessels dilate to increase heat loss |
|
Sweat glands |
None |
Produce sweat; evaporation increases heat loss |
|
Brain |
Behavioral (nonphysiological) responses-- put on coat, curl up, etc. |
Behavioral (nonphysiological) responses -- take off coat, etc. |
4. Cooling vs. Heating -- What can effectors do? Effectors can increase or decrease heat loss; can only increase heat generation. (Cannot decrease heat generation.) Therefore ability of humans to cope with very cold environments is better than their ability to cope with excessively hot environments.
5. Does drinking make you warmer in winter? See this 'Really?' column from the NYTimes.
Try Problem 5-2, c. & 5-5.
D. Body Temperature and the General Case -- The Circuit View -- handout 15A, top.
1. Circuit = 1 loop of seesaw. Seesaw = double circuit. Often two circuits to make opposite types of corrections.
2. Signals: Signals can be hormonal or neuronal.
3. Afferent vs Efferent Signals. Bottom half of circuit has two arms -- afferent vs efferent
Afferent information goes from sensors → in to IC
Efferent goes out of IC → toward effectors
4. Regulation vs Control.
a. Regulation/regulated variable: The variable (glucose level) you wish to keep at an approximately constant level is said to be "regulated."
b. Control/controlled process: The processes that alter levels of the regulated variable (glucose uptake, release or shivering, sweating, etc.) are said to be "controlled."
c. What's the difference?
The point of the system is to maintain homeostasis of blood glucose levels, internal temperature, etc. Not to maintain homeostasis of rates of glucose uptake, sweating, etc.
The value of the regulated variable stays about the same; the rates of the controlled processes (glucose uptake, heat loss, heat generation, etc. ) can vary as much as necessary to achieve homeostasis of blood glucose levels or temperature.
5. May be multiple effectors and/or sensors.
6. IC (when there are multiple inputs) is nervous tissue or brain.
a. Major Role -- Compares current value to set point; sends appropriate message to effectors.
b. Adjustments -- IC can adjust set points and/or critical points. Why bother? Fevers & feedforward:
(1). Fevers -- Raise set point for body temperature and critical points for shivering/sweating
Shivering and sweating both kick in at higher temps. (You don't have to cool off as much to start shivering and you need to heat up more to start sweating.) Raises set point (desired level) & actual level of internal body temperature.
Why fevers? High temperatures prevent bacteria from obtaining iron from host & improve immune function.
(2). Feedforward or anticipation -- Planning ahead. Altering set points and/or critical points to adjust to anticipated factors. (Or you can think of it as just ignoring the usual critical points.) Examples:
Body temperature: Skin temperature affects critical temperature/set points for generating heat and/or shivering. If body is cold, but it's warm outside, shivering can be postponed, saving energy, and you'll still warm up. This is equivalent to lowering (or ignoring) set point/critical points for shivering, not changing set point of internal body temperature. Changes what effectors and what controlled processes you use to warm up, but not the end result.
Secreting insulin when you start to digest food in the stomach, but before the digestion products (glucose, amino acids etc.) reach the blood. This way tissues will be ready to take up the glucose as soon as it enters the blood.
E. What other components of internal milieu are regulated besides glucose, temperature? Many nutrients like amino acids; concentrations of water, salts and ions (Na+, K+ etc.), gases (CO2, O2), waste products, volume & pressure of blood, and pH.
Try Problems 5-3, 5-4 & 5-9
A & B, & 5-10.
Next Time: Wrap up of homeostasis -- whatever we don't get to above, and
then how the effectors work.
{kind=link}