C2006/F2402 '11 -- Outline For Lecture #6
(c) 2011
Dr. Deborah Mowshowitz , Columbia University, New York, NY. Last update 02/06/2011 02:55 PMHandouts:
6A-- Transport of glucose through body (gif)
6A-- pdf
6B -- RME
(gif)
6B --RME (pdf)
6C -- Structure of Capillaries & Transcytosis (Posted
on Courseworks).
Here are links for a
diagram of a capillary, a
diagram of transcytosis,
and an
electron micrograph of a capillary.
I. Putting all the Methods of Transport of Small Molecules Together or What Good is All This?
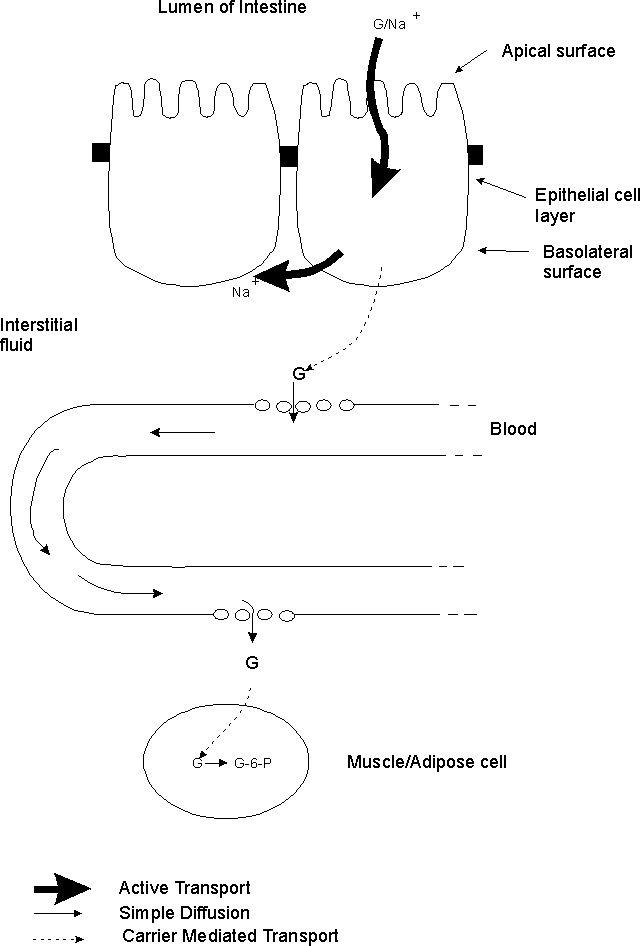
A. How glucose gets from lumen of intestine → muscle and adipose cells. An example of how the various types of transport are used. (Handout 6A) Steps in the process:
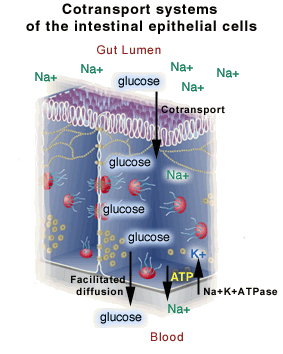
1. How glucose exits lumen. Glucose crosses apical surface of epithelial cells primarily by Na+/Glucose co-transport. (2o act. transport).
2. Role of Na+/K+ pump. Pump in basolateral (BL) surface keeps Na+ in cell low, so Na+ gradient favors entry of Na+. (1o act. transport)
3. How glucose exits epithelial cells.
a. Glucose (except that used for metabolism of epithelial cell) exits BL surface of cell by facilitated diffusion = carrier mediated transport.
b. Transporter protein = GLUT2 (more details on GLUT family of proteins below).
c. When glucose leaves cells it enters the interstitial fluid = IF = fluid in between body cells.
4. How glucose enters and leaves capillaries -- by simple diffusion through spaces between the cells. Cells surrounding capillaries in most of body are not joined by tight junctions.
a. Material does NOT enter capillaries by diffusion across a membrane. Material diffuses through liquid in spaces (pores) between the cells.
b. For structure of capillaries, see handout 6C, bottom. (Also see links at start of lecture.)Pictures are provided on handout since function is hard to understand without the anatomy. Picture shows how endothelial cells surround capillary lumen, forming pores between cells. Pores allow diffusion (of glucose and other small hydrophilic molecules, but not proteins) in and out of capillary.
c. Blood brain barrier: Capillary cells in brain are joined by tight junctions -- there are no pores (spaces between cells) so material cannot diffuse in and out of capillaries in brain. Click here for details on the BBB. (fyi only).
5. How glucose enters body cells
a. Glucose enters cells by facilitated diffusion = carrier mediated transport using a GLUT protein.
b. Carrier is permanently in cell membrane in many cell types (brain, liver). See below on GLUT transporters.
c. Carrier (GLUT 4) is only "mobilized" that is, inserted into membrane (by fusion of vesicles as explained previously) in some cell types (adipose & muscle) in presence of insulin.
6. Role of glucose phosphorylation. Conversion of G → G-6-phosphate traps G inside cells.
For additional examples of the uses of the various types of transport processes, see Becker fig. 8-1 & 8-2. For pictures of steps 1-3, see http://www.biology.arizona.edu/cell_bio/problem_sets/membranes/graphics/cotransport_sys.gif or
http://www.biochem.arizona.edu/classes/bioc462/462a/NOTES/LIPIDS/Fig12_36GlcNaSymport.GIF
Note both of these come from classes with extensive on line notes. The biochem course includes several animations of transport proteins.
B. How Glucose Reaches Body Cells -- Another look at handout 6-A. The steps in the process are described above in the order in which they occur. Here is a summary with the focus on the various types of transport involved.
1. Role of Active transport -- Needed to get glucose from lumen to inside of epithelial cell.
a. Primary active transport -- Na+/K+ pump keeps intracellular [Na+] low.
b. Secondary active transport -- Glucose enters epithelial cells by Na+/Glucose co-transport
2. Role of Passive Transport & Phosphorylation (of glucose)
a. Passive Transport -- Used to move glucose the rest of the way -- out of epithelial cells, in & out of capillaries, and into body cells.
b. Phosphorylation of glucose -- Used in the body cells to keep the free glucose level at the "end of the road" low, and ensure that the glucose gradient is "downhill" from epithelial cells to capillaries to body cells.
3. Role of Diffusion: Glucose and other small molecules (but not macromolecules) diffuse in and out of capillaries through the liquid filled spaces between the cells, not by diffusing across the cell membrane.
Most proteins are too big to enter or leave capillaries by diffusion. Most proteins enter and leave by transcytosis; shown on handout 6C and explained below.
4. Role of Channels: None are shown on the handout, but glucose can pass from one epithelial cell to another through gap junctions.
5. Role of GLUT transporters (another protein/gene family)
a. GLUT proteins are responsible for passive transport of glucose. All passive glucose transport across membranes (that is carrier mediated) depends on a family of proteins called GLUT 1, GLUT 2, etc. (GLUT = Glucose transporters)
b. Different family members (genes and proteins) are expressed in different cell types. GLUT 1 protein is found in plasma membrane of RBC & most other cells, GLUT 2 protein on BL surface of intestinal epithelial cells, GLUT 4 protein in muscle and adipose tissue, etc. (Note all genes for all proteins are present in all these cell types -- DNA is the same!)
c. All the genes and corresponding proteins are similar, but have significant structural and functional differences. This is another example of a gene/protein family. All the proteins have a similar overall structure -- 12 transmembrane segments, COOH and amino ends on intracellular side of membrane, etc. For a picture click here.; For a diagram and table go to http://www.ncbi.nlm.nih.gov/books/NBK30/box/A54/
d. Position & Action of GLUT 4 is insulin dependent. GLUT 4 is the only insulin dependent member of the family. Insulin triggers insertion of GLUT 4 protein into the plasma membrane, by triggering vesicle fusion, as explained previously. All the other proteins are located constitutively in their respective membranes.
e. Direction of transport. Note that one member of this family (GLUT 2) is responsible for ferrying glucose OUT of epithelial cells; different members are responsible for helping glucose ENTER most other cells. All family members bind glucose on one side of the membrane, change conformation and release glucose on the other side of the membrane. Which way the glucose goes (in or out) depends on the relative concentrations of glucose on the two sides of the respective membrane, not on which GLUT protein is used. (See problem 2-12 C.)
f. SGLT proteins are responsible for active transport of glucose. (SGLT = Sodium -glucose transporters). SGLT proteins make up a different protein family. The members of this family are responsible for active transport of glucose across membranes. (See problem 2R-2.)
g. All transport of glucose into and out of cells requires a transport protein. Protein can be a carrier, pump, or channel. Transport into capillaries by diffusion between the cells does not require a protein transporter.
Try problem 2-9 &
2-12.
II.
Ways that Big Molecules Enter
Cells -- Types of Endocytosis.
In all cases net effect is that cell membrane folds in and pinches off, forming
a vesicle in the cytoplasm that contains material from the outside.
A. Pinocytosis = bulk phase endocytosis; no receptor. Cells take in random samples of surrounding fluid containing a random selection of extracellular substances.
B. Phagocytosis -- in specialized cells only -- extensions of cells (pseudopods) reach out and engulf solids. See Becker fig. 12-14. Vesicle that is formed is called a phagocytic vesicle (or vacuole) or phagosome. Requires MF.
C. RME = receptor mediated endocytosis. Cells take in specific substances from surrounding fluid using a receptor. See Becker fig. 12-15 (diagram) & 12-16 (micrograph). Different cell types have different combinations of receptors.
III. RME -- Receptor Mediated Endocytosis
A. General and/or important Features.
1. Receptors -- Need specific receptor for each substance (or class of closely related substances) to be transported. 2. Concentrates substances transported -- usually moves them up their gradient. 3. Requires energy -- multiple stages in process use ATP or GTP. Energy must be required because substances move against their gradients. Energy is required to form the vesicles and to process and/or transport the vesicles inside the cell.
4. Role of clathrin -- A peripheral membrane protein is needed to deform membrane and allow vesicles to form -- provides a coat. (See Becker figs. 12-15 to 2-18 and/or Sadava fig. 6.19 (5.17)
Other proteins are required as well, but will not be discussed.a. Clathrin is coat protein for vesicles forming from plasma membrane and trans-Golgi*.
b. Budding of other membranes involve different "coat" proteins. Best known are COPI & COPII which are involved in ER-Golgi transport. (See Becker for details if interested. Types of coats are summarized in table 12-2.)
*Note on terminology: Trans side of Golgi = "far end" = side away from nucleus and ER = last part that proteins travel through as they are processed in Golgi. Also called "TGN" for "trans Golgi Network." See Sadava fig. 5.11 (4.11) or Becker fig. 12-8 for labeled pictures. More details on structure and function of Golgi later.
5. It's a cycle -- Exocytosis balances endocytosis so cell surface area stays the same. See Sadava fig. 6.18 (5.16) or Becker fig. 12-15. For LDL receptor, it takes about 10-20 minutes for one "round trip."
6. Topology -- material can enter and/or exit cell without being in contact with cytoplasm. Material can remain inside a vesicle or outside cell at all times. (See Transcytosis.)
7. Possible fates of endocytosed material -- Where does vesicle go? Where do receptor &/or ligand end up?
a. Degradation -- vesicle fuses with lysosomes and contents are degraded.
b. Return to surface -- material recycles -- vesicle fuses with plasma membrane.
c. Sorting -- not everything in the vesicle may go to the same place. Vesicle may fuse with endosome (sorting vesicle), and different parts of the endocytosed material may be directed to different destinations. (More details on a-c below.)
d. Transcytosis -- vesicle crosses cell and fuses with opposite cell surface. For examples see handout 6C or diagram of transcytosis (shows how antibodies enter lumen)
(1). Requires Receptor: Transcytosis requires a receptor for each substance transported. Receptor is not shown on 6C but is clearly shown on diagram of transcytosis
(2). Transcytosis = endocytosis + exocytosis = 2 steps
(a). Material binds to receptor and is endocytosed on one surface of the cell.
(b). Vesicle moves across cell and material is exocytosed on a different surface.
(3). Function. Can be used to move proteins, across a cell, in either direction. See examples above or RP3.
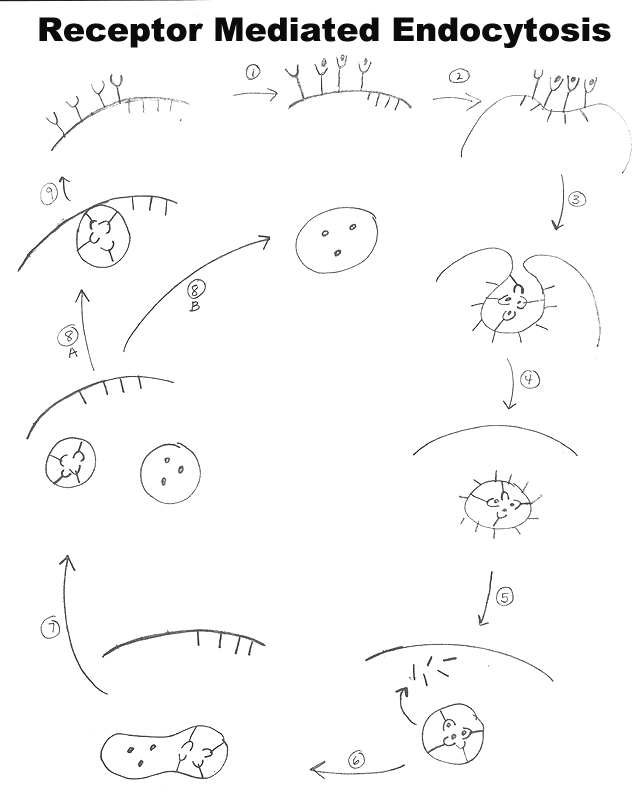
B. Stages of Cycle (Numbers match steps on handout 6B.) Click here for animation.
(1). Receptors bind material
(ligand) to be internalized(2). Receptors are in (or migrate to) coated pits = clathrin
-coated parts of membrane(3). Membrane starts to invaginate to form coated vesicle
. A single vesicle can contain more than one type of receptor plus ligand.(4). Coated vesicle forms
(pinching off of vesicle is an energy requiring step)(5). Uncoating occurs relatively quickly
(uncoating requires energy)(6). Vesicle is acidified to become endosome (or fuses with pre-existing endosome), and sorting of receptor(s) and ligand(s) begins.
- A single endosome may contain many different receptors and ligands, and different ones are sorted differently. (Some examples are given in detail below.)
- The uncoated, acidified vesicle can be called an endosome, early endosome, or a sorting vesicle.
- Acidification requires energy to run proton pump -- to move H+ into vesicle at expense of ATP. Pump is in membrane of vesicle.
Note: Details of sorting and recycling -- the remaining steps -- vary with material endocytosed. More details below for individual cases.
(7). Endosome splits
. The substance we are following, and/or its receptor, can end up in either half.
In example shown on handout, one half gets the receptor and one half gets the ligand, as is the case for LDL. Other examples will be discussed in class and are outlined in detail below.Note: endosome may not simply split in one step; process of sorting may be gradual. Pieces of different composition may gradually bud off as internal composition of remainder changes.
(8). What Happens to the Different Parts of the Endosome?
8A. Fate of vesicle with materials to be recycled (receptors and/or carriers) -- this vesicle fuses with plasma membrane in step 9. (In case of LDL, this vesicle would contain the receptor for LDL.)
8B. Fate of vesicle with material that remains inside the cell -- Vesicle delivers contents to appropriate cell compartment. (For LDL, vesicle delivers LDL to lysosomes, so material is degraded.)
(9). Exocytosis occurs
-- returns receptors and/or other components to the plasma membrane or outside of cell.
Try Problem 2-6.
C. Some Specific Examples
1. LDL (Low density lipoprotein) -- receptor recycled, but ligand (including protein part) degraded. See Becker, Box 12B or text of Sadava Ch. 51.4 (50.4). Many of LDL details may have been included in general case, but are summarized below. Click here for a picture of LDL.a. What is LDL? A lipoprotein particle containing cholesterol esters + some other lipids + a protein. Particle contains esterified cholesterol covered by monolayer of amphipathic lipid (phospholipid plus some unesterified cholesterol) + one molecule of protein (apoprotein B or apoB).
2. EGF (Epidermal Growth Factor) -- all the protein involved (ligand + receptor) is degradedb. Why LDL?
(1) Why a monolayer on outside? Solubility. Cholesterol is insoluble in blood. (Too hydrophobic.) Need a way to ferry cholesterol through blood and into cell -- Cholesterol transport requires formation of particle with hydrophilic surface
(2) Why a protein (apoB)? For binding to cell surface receptor (LDL receptor). A protein is needed as ligand to bind to receptor.
(3) Summary of Roles of Parts of LDL:
(1). Protein (apoB) = Ligand = what actually binds LDL receptor = protein part of LDL
(2). Cholesterol -- What cell actually needs is the cholesterol part (for building its membranes &/or hormone synthesis).
c. Receptor, but not protein part of LDL, is recycled. Note: there are 2 separate proteins here that are easily confused
(1) Receptor protein on the cell surface = LDL receptor = binds LDL and allows uptake of cholesterol
(2) Protein in LDL (apoB) = ligand for LDL receptor = part of LDL and helps carry cholesterol through the blood.
d. Receptor and apoB are separated inside sorting vesicles/endosomes. All of LDL (including protein) stays together; separates from receptor
e. Need lysosomes to degrade LDL protein and release cholesterol (cholesterol esters in LDL must be split for cholesterol to be used). How does LDL reach lysosomes? Through fusion of vesicles. Either:
(1). Vesicles/endosomes holding substrate fuse with pre-existing lysosomes, or
(2). Vesicles with substrate fuse with vesicles from Golgi carrying newly made hydrolases to form new lysosomes. (More details on how hydrolases pass through the Golgi and are targeted to lysosomes to be discussed later.)
f. Function of LDL uptake -- to supply a nutrient (cholesterol).
g. Current terminology: relationship of early endosomes, late endosomes & lysosomes. Note: Most of this is FYI. In this course, the term "endosomes" will be used for both early and late endosomes.
(1). Early endosome = sorting vesicle. Term is used differently by different authors. Can be "early" on pathway into cell (by endocytosis) and/or "early" on pathway from Golgi to lysosomes. Therefore, early endosomes can mean:
(a) Uncoated & acidified vesicles from invagination of plasma membrane carrying newly endocytosed material,
(b). Vesicles coming from Golgi carrying newly made proteins (more on this later).
(2). Late endosome = vesicle containing hydrolytic enzymes destined for lysosomes (but not yet activated) plus potential substrate. More acidic than early endosome. Material not destined for lysosomes has been jettisoned. Formed by maturation of early endosome.
(3). Lysosomes = vesicle containing active hydrolytic enzymes and substrate. More acidic than late endosome. Formed by maturation of late endosome and/or fusion with pre-existing lysosome.
(4). Older terminology found in some texts (FYI only):
(a). Primary lysosome = vesicle with hydrolytic enzymes only.
(b). Secondary lysosome = enzymes + substrate = result of fusion of primary lyso. + another vesicle containing substrate.
a. No separate protein ligand required; EGF is a protein -- unlike cholesterol, or Fe (see case below). EGF itself binds to receptor = ligand for cell surface receptor & substance that will be transported into the cell.
b. Function of uptake -- to regulate signaling. EGF is a signaling molecule. Uptake turns off signal and down regulates receptors (reduces # of cell surface receptors).
c. Receptor not recycled -- Ligand (signal molecule) and receptor degraded together.
d. Need lysosomes (to degrade both receptor and ligand).
3. Fe/Transferrin -- none of the protein involved is degraded -- all recycled
a. What is transferrin? Fe needs a protein (like cholesterol needs apoB) for transport and binding to receptor; protein (= ligand for cell receptor) is called transferrin.
b. Both apotransferrin & receptor are recycled.
c. No lysosomes needed -- iron is transported out of endosome (using transporter protein or channel in membrane); no protein is degraded.
d. Transferrin and receptor separate outside cell after recycled
(1). Fe/transferrin binds to receptor at neutral pH and enters cell by RME.
(2). Inside cell, Fe transported out of vesicle into cytoplasm, leaving apo-transferrin stuck to receptor ("apo" means without ligand, cofactor, etc.).
(3). Apo-transferrin (= transferrin without Fe) sticks to receptor at low pH (in endosome) but separates at neutral pH (outside cell). This is contrary to usual behavior -- Most ligands stick to receptors at neutral pH but separate at low pH found in endosome. (Low pH breaks many weak bonds.)
(4). Note that apo-transferrin and Fe/transferrin have different affinities for the receptor at neutral pH. Under these conditions (neutral pH), Fe/transferrin binds to the receptor, and apo-transferrin separates from the receptor.
e. Function of uptake -- to supply a nutrient (Fe).
D. For Reference: Compare & Contrast for the examples described above for transport of X
| Transferrin | LDL | EGF | |
| What's carried in (what is X)? | Fe | Cholesterol | Growth Factor |
| Function of X | Metabolism (Fe is cofactor for many proteins) | Metabolism (cholesterol is a component of cell membranes; used for hormone synthesis) | Signal |
| Ligand (What binds to receptor?) | Transferrin = apotranferrin + Fe | LDL | EGF |
| Does ligand include protein in addition to X? | Yes (apotransferrin) | Yes (ApoB) | No |
| Ligand Fate (protein part) | Recycled | Digested | Digested |
| Receptor Fate | Recycled | Recycled | Digested |
| Do protein part of ligand & receptor separate inside cell? | No | Yes | No |
| Lysosomes Involved? | No | Yes | Yes |
| Where do ligand & receptor separate? | Outside the cell | In endosomes | Not separated -- both degraded |
A. Types of Labeling (using added tracers)
B. Detection -- How do you find where the radioactivity (or whatever tracer/label you used) is?1. Continuous Labeling -- switch from regular, ordinary material to labeled material (material containing radioactivity, fluorescence, etc.) and follow what gets labeled first (with radioactivity, fluorescence, etc.), what gets labeled next, and so on. We will discuss radioactive labeling, but the principle is the same whether the label is radioactivity, fluorescence, etc.
2. Pulse-Chase Experiments -- supply radioactive material for a brief time (pulse) and then switch back to ordinary, non-radioactive material (chase). Follow where the radioactivity goes. The "pulse" passes through the cell like a mouse through a boa constrictor. Just as different parts of the boa constrictor bulge out temporarily as the mouse passes down the snake, so different parts of the cell become radioactive temporarily, one at a time, as the radioactive material passes through. Then as the "pulse" or the "mouse" passes on, each part will return to normal -- non radioactive or normal size, depending on whether we are referring to the cell or to the snake.
1. Autoradiography -- Cover a layer of labeled cells with photographic emulsion and count radioactive grains over each organelle or part of the cell. This method is similar to doing in situ assays, in that you examine intact cells to pin down the location of what you are looking for. See Becker, Appendix, A-17 (A-18) or Guide to Microscopy.
Note: Becker's Appendix (or Guide to Microscopy in 5th ed.) has a lot of useful background info on microscopic methods, including immunofluorescence, freeze fracture, etc.
For a picture of typical results, see fig. 12-10 of Becker (7th ed) or click here. These pictures were obtained by following newly made material out of the cell, not following it in. However the principle is the same -- it shows label in different cell parts at different times.
2. Fractionate First -- Break up labeled samples, fractionate into various organelles, and measure radioactivity in each fraction. This is similar to the "grind and find" procedure, in that you break up the cells, separate them into their parts, and test a solution or suspension of each part for what you are looking for.
To review labeling and RME, try problems 2-8 & 2-11; by now you should be able to do all the problems in problem set 2 & 2R.
Next Time: How do proteins get sorted to their proper place? How do molecules get in and out of the nucleus?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}