C2006/F2402 '11 OUTLINE OF LECTURE #7
(c) 2011 Dr. Deborah Mowshowitz, Columbia University, New York, NY. Last update 02/08/2011 09:56 PM
Handouts :
7A -- Sorting of Proteins (Overview).
7B -- Types of Signaling
NY Times Sunday Magazine of 2/22/09 has an article that you might find interesting about the case of a woman who may have a defect in her 'basement membranes.' Article is really about how doctors try to diagnose mystifying conditions.
I. Labeling -- How do you follow material into or out of the cell? (Handout 6C; see notes of last time for details of A & B )
A. Types of Labeling -- Continuous vs Pulse-Chase
For a nice video (& explanation) of a pulse
chase experiment, try this link:
http://www.sumanasinc.com/webcontent/anisamples/majorsbiology/pulsechase/pulsechase.html
This site has many interesting animations. To select a topic, go to
http://sumanasinc.com/webcontent/animation.html
B. Types of Detection -- Autoradiography (in situ cell bio method) vs Fractionation (in vitro biochemical method)
C. Labeling -- How do you follow newly made molecules moving through the cell and/or on their way out? How do we know newly made proteins go from RER to Golgi etc.?
In examples of detection discussed previously, emphasis was on following molecules going in to the cell. This example is about following newly made molecules on their way out.
-- Add labeled precursors (small molecules) and measure incorporation into macromolecules.1. General idea
a. Add labeled precursors, and take cell samples after increasing time intervals.
b. For each sample, wash out unused ('unincorporated') small molecules -- removes labeled molecules not used for synthesis so not incorporated into macromolecules. Radioactivity remaining in dif. parts of the cell is in macromolecules.
c. Use autoradiography for measurement of radioactivity in each cell part, or measure amounts in each isolated fraction.
2. A specific example
-- following secreted proteins out. See graphs on handout 6C. Re-label the curves, left to right, with Rough ER, Golgi, Vesicles, Outside the cell.a. Continuous label vs pulse-chase results (See graphs on handout 6C & fig. 12-10 of Becker. 6th ed. has curves; 7th ed. has autoradiographs.)
b. Implications: newly made proteins to be secreted go → RER → Golgi → secretory vesicles → outside (See Becker fig. 12-8.) Click here for animation.
To review labeling of newly made material, try 3-4D.
D. Another Type of Labeling -- Cell makes its own labeled (fluorescent) protein containing GFP
GFP has been mentioned before. Here are the details.
1. What is it? GFP = green fluorescent protein = small fluorescent protein made by jelly fish. (Click here for page with pictures of GFP and related fluorescent proteins.)
2. What is it good for? GFP is used as tag to follow proteins inside the cell.
3. How is GFP added to proteins? GFP is not added from outside. Instead, genetic engineering is used to splice the gene for GFP to the gene for the protein of interest. The recombinant gene makes a fusion protein = normal sequence of amino acid + sequence of amino acids in GFP. Fusion protein (including GFP) is made internally by the cell; the functioning protein part and the GFP part fold up separately (each forms a separate domain).
4. How does fusion protein work?
a. GFP part fluoresces. In other words, cell makes its own fluorescently tagged version of the protein.
b. Functioning protein part usually works normally, but location of protein can be easily followed in cell, because protein has GFP attached. GFP labeled protein is used for many purposes, including following newly made protein through the cell.
5. Examples. For examples of use of GFP, see Becker fig. A-14, or Sadava fig. 18.5 9th ed. (Description, but no picture in 8th.)
GFP is often used to identify cells that express (turn on) a particular gene. For an example see this picture. The cells that "light up" are the only ones that express (turn on) the fusion gene. Only these cells produce a fusion protein containing GFP. (This example also illustrates why people use small, transparent organisms as "model organisms.") For a really startling picture, and article try this link.
See also: http://nobelprize.org/nobel_prizes/chemistry/laureates/2008/presentation-speech.html.
See problem 2R-4 for an example of the use of GFP labeling.
Note: Material through here will be included in Exam #1. All remaining material will be covered in Exam #2.
II. Sorting of Proteins to their Proper Place: Overview See Handout 7A, or Becker fig. 22-14 (20-14) or Sadava 14.19 (12.15) -- terminology in Sadava is slightly different.
A. What determines the fate (final location) of each individual protein? The amino acid sequence of the protein itself. The ability of each protein to reach its proper destination is built into the protein. The presence (or absence) of localization signals in the amino acid sequence of the protein is the determining factor.
1. What's a localization signal: a group of amino acids acting as an "address" or "tag" directing the protein to a particular destination.
2. Terminology: The localization signal or "tag" is often called a localization sequence (LS) or patch.
a. LS -- if it consists of a continuous section in the peptide chain.
b. Patch -- if it consists of a contiguous section in the folded protein. (But AA are not next to each other in the unfolded chain.)
3. Use of tags: The localization sequence/patch directs the protein to the ER, nucleus, etc. Several different localization sequences, which are read sequentially, may be needed to direct a protein to its proper destination. If no "tag" or special sequence/patch is present at all, the protein remains in the (soluble) cytoplasm.
B. The Big Divide -- Attach to the ER or not?
1. Ribosomes start to make protein first. Translation starts first; then ribosome location (attachment to ER or not) is determined by the sequence of the protein being made.
2. Which ribosomes go to ER: If protein has the right "tag" (a localization signal called a signal peptide (SP) or signal sequence) -- the ribosomes attach to the ER, and protein enters ER as it is made.
3. Which ribosomes stay in cytoplasm: If there is no SP, ribosomes remain "free" in the cytoplasm -- they do not attach to ER or any other membrane.
C. Fate of proteins made on free ribosomes
1. Soluble Cytoplasm -- the default location for a soluble protein. If there are no "tags" at all, proteins stay in the cytoplasm.
2. Organelles. If proteins have the right "tags" they can be imported post-translationally (after synthesis) into organelles (nuclei, mito, chloro or peroxisomes) that are NOT part of the endomembrane system.
3. Terminology: "free" means not attached to a membrane. All ribosomes making protein, "free" or not, are attached to mRNA.
D. Fate of proteins made on attached ribosomes -- these become part of the endomembrane system and/or leave the cell. How they enter the ER & other parts of the EMS will be discussed in more detail in later lectures.
1. They enter the ER as they are made (co-translational import). Protein can
- Remain inserted in membrane -- become a transmembrane single or multipass protein (details later)
- Pass all the way across the membrane and end up in lumen of ER (as soluble protein or peripheral, membrane associated protein on lumen side)
2. Most proteins travel from ER to Golgi (a few may remain in ER).
3. Where do proteins go when they leave the Golgi?
b. To the plasma membrane -- Result is release of contents of vesicle outside cell and/or addition of material to cell membrane. Click here for animation #1 -- annotated & animation #2 -- larger but not annotated.
4. How do proteins travel from one part of EMS to another or to the plasma membrane? Vesicles carrying protein bud off ER, Golgi, etc., travel to another compartment (or plasma membrane) and fuse with target membrane.
a. Transmembrane proteins remain in the membrane. TM proteins move from membrane to membrane but always remain in the same orientation -- cytoplasmic domain remains in cytoplasm.
b. Soluble proteins stay soluble. Contents of lumen of compartment #1 → lumen of vesicle → lumen of next compartment or outside the cell.
Questions (to check your understanding of the
topology):
(1). Suppose domain X of a multipass integral membrane protein is on the
inside of the ER, sticking into the lumen of the ER. When a vesicle forms
off the ER, where will domain X be? Inside vesicle in lumen? On outside of
vesicle in cytoplasm?
(2). When vesicle fuses with Golgi, where will domain X be? In lumen of Golgi? Sticking out into cytoplasm?
III. Extracellular Proteins & Signaling -- what are extracellular proteins good for?
A. Important types of secreted Proteins
1. Components of ECM. Examples: collagen, fibronectin, laminin, elastin.
2. Signaling molecules -- endocrines, paracrines & autocrines -- secreted into IF or blood stream. Examples: insulin, growth hormone, glucagon, oxytocin, prolactin.
3. Exocrines -- secreted to outside of body or inside of lumen (such as lumen of GI tract). Examples: Pancreatic enzymes (Trypsin, chymotrypsin) & proteins of breast milk.
B. Why do you need signaling molecules?
1. For development to work properly
2. To co-ordinate functions in adult -- between cells, tissues and organs (w/o using nerves)
3. For nerve-nerve and nerve-target communication -- most of this involves small molecules as signals and proteins as receptors. Will be discussed when we get to nerves.
C. Major types of secreted Signals -- classified here by type of cell that makes them and/or target location. See Handout 7A for pictures -- numbers of pictures match numbers below. Many, but not all of these are peptides.
1. Endocrine:
a. Signal molecule secreted by specialized cells in ductless (endocrine) gland
b. Gland secretes signal molecule (hormone) into blood.
c. Target cell is often far away. Acts long range. For an example see Becker fig. 14-23 (14-22).
d. Examples of proteins that act as endocrines: Insulin,
e. Endocrine is not always a protein or peptide -- can be a protein (insulin & many others) or not (testosterone or other steroid or thyroid hormone)
2. Paracrine: See Becker fig. 14-1 & table 14-4 (6th ed) for paracrine (or autocrine) vs. endocrine.
a. Usually secreted by ordinary cells
b. Target cell is near by -- Receptor is on adjacent cells. Act locally.
c. Example -- many growth factors (such as EGF); Wnt proteins (products of WNT genes), AMH (AntiMullerian Hormone -- not an endocrine, in spite of name)
For use of PRP (Platelet rich plasma) to supply GF's acting as paracrines see http://web.jbjs.org.uk/cgi/content/full/91-B/8/987
3. Autocrine: Like paracrine, except receptor is on same cell. ex. = some growth factors
4. Neurocrine:
a. Neuron secretes signal molecule.
b. Signal molecule acts as a neurotransmitter (NT)
c. NT acts on receptors on neighbor (gland, another neuron or muscle). Acts locally, like a paracrine.
d. Examples: norephinephrine, acetyl choline. Details when we get to nerves.
e. Most neurocrines are small molecules, not proteins. (A few are short peptides.)
5. Neuroendocrine:
a. Neuron secretes signal molecule, as in previous case.
b. Signal molecule acts like a hormone (travels through blood to target).
c. Example: Epinephrine (adrenaline).
6. Exocrine:
a. Exocrine gland secretions are released by ducts to outside of body (or to inside of body cavity).
b. Typical Exocrine secretions
(1). Released to outside: sweat, tears, breast milk
(2). Released to spaces inside of the body that connect with the outside, such as the GI lumen: enzymes of pancreas.
c. Secretions on outside can carry signals → target in different individual = pheromones (detected by olfactory receptors in mammals).
D. Other types of Signaling
1. Gap Junctions -- allow ions & currents to flow directly from cell to cell -- used in smooth muscle → synchronized contractions. Sadava fig. 7.21 (15.19).
2. Juxtacrine. Cell surface proteins from two different cells contact -- used in immune system. Similar to basic system, but signal molecule is not secreted -- remains on cell surface.
IV. How
Proteins Enter Nuclei -- Nuclear Pores and Traffic through Them
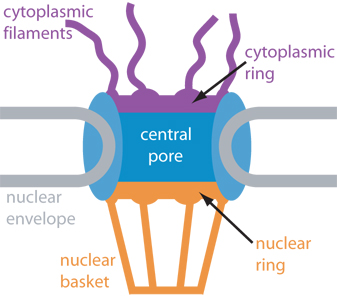
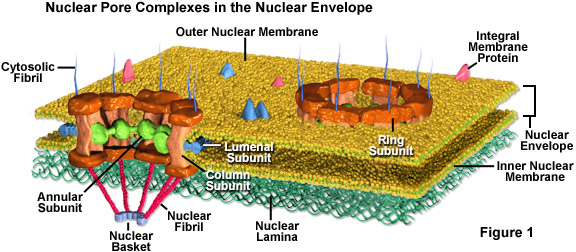
A. Pore structure -- Large complex structure that is hard to explain; will be demonstrated. See Becker fig. 18-27 & 28 and/or Sadava fig. 5.8 (4.8). Some links to pictures are below. Go to Google images for more pictures.
Two depictions of structure of a nuclear pore: Diagram #1; Diagram #2.
Diagram of how pores are located in the membrane.
Freeze fracture showing pores on surface of nucleus.
B. Pore Function
1. For relatively small molecules: Acts like (ungated, always open) channel for small molecules (may include small proteins). Transport is passive.
2. For big molecules: Acts as active transporter for big molecules. Features of active transport are below.
3. Transport is bidirectional -- mRNA, ribosomal subunits etc., must go out (see below) and proteins (as well as small molecules) must go in. (See Becker fig. 18-29; molecular details are in 18-30.) How things get out is not as well understood as how things get in.
4. Important Features of Protein Transport through Nuclear Pores
a. Transport requires a localization signal. Protein to be transported requires NLS (nuclear localization signal) to be transported into nucleus; NES (nuclear exit signal) on protein to be transported out.
b. Transport requires energy (GTP split).
c. 'Middle-man' protein required. Transport requires importin (to go in) or exportin (to go out).
d. Transport involves binding to pore protein.
Protein with NLS binds to "middle man" or "ferry proteins" (importins or exportins) which bind to pore protein.
Importin goes into the nucleus with the 'cargo' protein it is transporting in; exportin goes out with its cargo; both proteins then recycle.
Additional proteins (that we will ignore) are required as well. See Becker fig. 18-30 for a model.
Mechanism of transport through nuclear pores is similar, but not identical, in both directions.
e. Soluble and peripheral proteins are transported through pores. How do integral (transmembrane) proteins of the nuclear membrane reach their destination? Not through pores. TM proteins are probably made on the ER, and slide laterally into the outer & inner nuclear membranes (continuous with the ER). Once in place, TM proteins are anchored by binding to lamins or other internal nuclear proteins. Details of how TM proteins are inserted in membranes will be covered later.
To review nuclear transport, try problem 4-2.
V. Nuclear Structure & Role of nucleolus. See handout 7B
A. Lamins A, B, C & Nuclear lamina
1. Lamins = IF's of nucleus (No MT or MF in nucleus)
2. Not to be confused with Laminin -- glycoprotein in the extracellular matrix (ECM) found primarily in the basal lamina
3. Form nuclear cortex or nuclear lamina = web supporting the nuclear envelope -- see Becker fig. 18-31(b). Similar function to ankryin/spectrin/actin web under plasma membrane -- provide support and points of attachment.
4. State of Phosphorylation and function change during cell cycle
a. Phosphorylated at G2 → M transition; Completely phosphorylated by prophase; due to rise in protein kinase activity; de-polymerize.
b. Dephosphorylated at anaphase, at start of M → G1 transition; due to rise in protein phosphatase activity; re-polymerize.
c. This is another example of using addition/removal of phosphate to control protein conformation & activity.
Kinases & phosphatases here are separate from their target proteins (as vs. case of Na+/K+ pump).
Same kinase or phosphatase can modify multiple target proteins at critical points in cell cycle. (Many proteins phosphorylated at entry to M and dephosphorylated at exit to G1.)
5. May help organize chromatin; may provide attachment sites for chromosomes and nuclear pore complexes.
Question: Should lamins have an NLS? NES? Should the LS be removed?
The material below will be covered in lecture #8.
B. Nucleolus
1. Nucleolus has no membrane (see Becker fig. 18-33).
2. Nucleolus is site of rRNA synthesis and assembly of ribosomal subunits
a. Production of ribosomes requires extensive traffic in and out of nucleus
All transport in and out of nucleus is through nuclear pores.
Both RNA and protein must pass through pores.
Reminder: importin goes into the nucleus with the 'cargo' protein it is transporting in; exportin goes out with its cargo; both proteins then recycle. (See Becker 18-30.)
b. rRNA is made in the nucleolus (see Becker fig. 18-34) using RNA polymerase I (see below)
c. Ribosomal proteins are NOT made in the nucleolus.**
Ribo proteins are made in the cytoplasm, but imported into the nucleus for assembly into subunits.
Messenger RNA for ribosomal proteins is made in the nucleus using polymerase II (see below)and exported to the cytoplasm, as for any other mRNA.
After the mRNA is translated, the proteins are imported back into the nucleus for assembly with rRNA.
**Note: Becker says there are ribosomes translating mRNA in the nucleus. Maybe, maybe not. This finding is controversial -- the existence of nuclear ribosomes is not firmly established, so we are ignoring them. Ribo proteins are made in the cytoplasm in any case.
d. Ribosomal subunits are assembled in the nucleolus from ribosomal proteins (made in cytoplasm) and rRNA.
e. Completed ribosomal subunits are shipped out to the cytoplasm to be used in protein synthesis.
4. Nucleolus vs NOR (= nucleolar organizer region of DNA)
a. There are multiple copies of genes for rRNA; many copies are located in tandem. There may be one or multiple clusters of rRNA genes. Each cluster = one NOR.
b. Why many copies of rRNA genes? Multiple copies are needed because rRNA is made directly off the DNA -- no intermediate such as mRNA to allow for amplification. (Q to think about: Do you need multiple copies of the genes for tRNA?)
c. NOR = region of the DNA coding for rRNA = rDNA = genes for rRNA. Each NOR has multiple copies of the rRNA genes. Genome may have one or more NORs. NORs are permanent -- are part of DNA.
d. Nucleolus = structure seen in interphase; not in meiosis/mitosis.
Nucleolus forms during interphase at location of one or more NORs.
Several NORs can cluster together to form one nucleolus.
Nucleoli are only visible when NOR is being transcribed and ribosomes are being assembled.
e. Nucleolus (but not NOR) disappears during meiosis/mitosis since genes for rRNA inactive then (no rRNA made or ribosomal subunits assembled)
C. RNA polymerases & associated proteins
1. In Prokaryotes -- there is only one RNA polymerase.
2. In eukaryotes -- there are three RNA polymerases. Differ in sensitivity to drugs, location in nucleus, & what RNA's they make. (See Becker table. 21-1 for details; for reference only.)
a. RNA pol I → big ribosomal RNAs. (All ribo RNA but 5S.) This is the polymerase found in the nucleolus.
b. RNA pol II → mRNA (We'll focus primarily on this one when we discuss regulation of transcription.)
c. RNA pol III → small RNA's like tRNA, 5S RNA (small ribosomal RNA)
3. Eukaryotic Transcription factors (TFs)
a. Eukaryotic transcription requires additional proteins (TFs), not just RNA polymerase.
b. TFs are needed to allow polymerase to bind to DNA
c. There are two types of TFs (more details next time)
(1). basal (or general) -- required for transcription of all genes; are the same in all cells
(2). regulatory -- decrease or increase basal transcription (usually called repressors or activators respectively). Are different in different cell types.
To review nuclear structure & transport into nuclei, try problems 4-2, 4-6 parts A & B, and 4-7, parts A & C.
D. Chromatin. DNA in eukaryotes is not "naked" as it is in bacteria. It is in the form of
chromatin = DNA complexed with proteins. So how do polymerases and TFs get to the DNA and how do they manage to carry
out transcription with all those proteins stuck to the DNA? What
actually turns transcription on and off? To answer
these questions, we need to look first at chromatin structure.
VI. General Chromatin Structure -- Euchromatin & Heterochromatin (Probably Next Time)
A. Composition of chromatin = DNA + associated proteins
1. Associated proteins are mostly histones (small, basic; more details to follow)
2. Associated proteins includes some (nonhistone) regulatory proteins
B. States of chromatin
1. In all states, DNA has proteins attached
2. Usually differences are due to different states of folding after histones added, not removal of histones (there are some exceptions which will be discussed later).
3. Two Types of Chromatin -- Two basic states of chromatin are visible in the light microscope
a. Heterochromatin or heterochromatic chromatin/DNA
(1). What is it? Chromatin that is darkly stained, & relatively condensed (tighter),
(2). Heterochromatin is genetically inactive (not transcribed)
(3). Two kinds of (interphase) heterochromatin
(a). Constitutive heterochromatin
Always heterochromatic in interphase in all cells
Example: chromatin at centromeres, telomeres
Usually repeating in sequence, non coding.
(b). Facultative heterochromatin
Sometimes heterochromatic in interphase, sometimes not (depends on tissue, time etc.).
Example: inactive X. Same X is not inactive in all cells.
Whichever X is inactive (forms a Barr body) is heterochromatic.
(4). Most of the DNA that is inactive during interphase (in a particular cell type) is NOT heterochromatic. Details next time.
(5). All chromatin is heterochromatic (tightly condensed) during mitosis & meiosis.
(6). All chromatin, both heterochromatin & euchromatin is replicated in S. Heterochromatic DNA is not genetically active (not transcribed), but it is replicated.
b. Euchromatin or euchromatic chromatin/DNA
(1). What is it? Chromatin that stains more lightly, is less condensed (looser).
(2). Euchromatin is capable of genetic activity (transcription) as vs. heterochromatin. Normal state of most DNA during interphase. Transcribable, but not necessarily being transcribed now.
(3). Most interphase DNA is euchromatic, whether it is transcribed or not. DNA must be euchromatic to be active, but not all euchromatin is active.
(4). Not all euchromatin is folded equally tightly
(a). Tighter vs looser. Euchromatin is often divided into several distinct states of folding, although tightness of folding is probably really continuous from relatively loose ↔ relatively tight.
(b). Different states of (eu)chromatin look about the same in the light microscope. Therefore indirect methods (such as DNase treatment) are necessary to test state of folding.
(c). Correlation between folding and function. In general, more active (transcribed) chromatin is looser, but the situation is complex .
Next Time: How are histones & DNA arranged in chromatin? Is looser chromatin more genetically active (more transcribed) and vice versa? How will you tell?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}