
(1) 26 pts. Answer TRUE or FALSE. If the statements are true, explain or give a supporting example. If the statements are false, either correct the statement or give a counter example.
a) Control by regulated transcription termination is necessarily unique to prokaryotes.
False. The form of regulated trx termination called "attenuation" requires coupled trx and translation which occurs only in prokaryotes. However, other forms to regulated trx termination could occur in eukaryotes. For example there is nothing to prevent a bgl operon-type mechanism from working in eukaryotes.
b) In humans, reverse transcriptase is found only in individuals infected with retroviruses.
False. Retroelements such as the ubiquitous LINES also encode reverse transcriptase.
c) i) Most oncogenic retroviruses require a helper virus to propagate.
True. Most RNA tumor viruses carry a modified version of a c-onc gene that has replaced essential viral functions.
ii) Oncogenic retroviruses carrying mutations in the env gene require a helper virus
to cause tumors.
False. An env mutant virus could infect a cell, convert its genome to DNA and integrate. Once integrated the v-onc gene could cause unregulated cell growth. The virus could not make more of itself without a helper but that's not the same thing as producing a cancer.
d) All genes that affect dorsal/ventral patterning in Drosophila are maternal effect genes.
False. While the genes that initially set up the Dorsal gradient are maternal, the targets of the Dorsal protein are expressed zygotically. The zen, twi, rho, and snail proteins are activated or repressed by Dorsal in the embryo--that's a zygotic effect.
e) In order for a two-hybrid system based on the lexA operator and DNA binding domain and the GAL4 activation domain to work, the yeast strain must contain a GAL80- mutation.
False. The presence of GAL80 protein would prevent the GAL4 protein from activating transcription under normal conditions. However the repression could simply be relieved by adding galactose to the growth medium. GAL80 is inactivated by galactose so that it will not repress GAL4 transcription activation function in the presence of glucose.
f) ES cells from a black (B/B) strain of mice are made mutant for one copy of the IRAK locus and injected into a blastocyst derived from brown (b/b) colored mice. If this blastocyst develops into a mouse and the mouse is crossed to a brown mouse, one-quarter of the progeny will be black-colored and heterozygous for the IRAK mutation.
False. The proportion of transgenic progeny depends on whether the germline was transformed. Chimeric mice whose germ cells derived from the b/b host would never produce progeny with the IRAK mutation. Chimeric mice that had some germ cells derived from the B/B IRAK/+ donor cells would produce some IRAK/+ heterozygotes but the proportion will vary depending on how many germ cells derived from the IRAK mutant cells.
2) 24 pts. Short answer section.
(a) Offer an explanation for why retro-elements generally account for a larger fraction of the genome than do transposable elements akin to P elements or the Ac/Ds elements of maize.
When a retroelement moves it always leaves a copy of itself behind in the genome. P elements and their relatives excise when they move to a new location so that the original transposon is often lost.
(b) Cancers can be induced by exposure to certain chemicals. Most such carcinogens are mutagens, however, some chemical carcinogens seem to cause cancer without inducing mutations. Briefly offer explanations for why mutagens may induce tumors and for how non-mutagenic carcinogens might act.
Mutagens can induce DNA changes that can inactivate tumor suppressor genes or activate oncognes resulting in uncontrolled cell growth. Non-mutagenic carcinogens could stimulating cell growth by directly affecting the proteins or cellular signals controlling growth.
(c) Partial diploids of E. coli that are IS/I+ are uninducible, whereas I-d/I+ partial diploids are constitutive. What do you think would be the phenotype of an IS/I-d strain? Explain your answer.
The IS/I-d strain would be constitutive. IS tetramers cannot bind inducer and thus repression cannot be relived by adding inducer. I-d mutations lock the repressor monomers into the induced non-binding state. Since even one I-d subunit poisons a tetramer whether the other subunits are wt or lacIS the I-d phenotype wins out.
(d). Draw the result of the PaJaMo experiment in the left graph. On the right graph, draw the predicted curve if the donor strain carried a lacIQ mutation. Genotypes are identical expect for the indicated difference. Briefly explain your answers.
In the PaJaMo experiment B-gal synthesis was initiated upon transfer into the recipient but ceased once sufficient repressor accumulated to repress the operon. Addition of IPTG restored lacZ synthesis demonstrating that the repressor was controlling expression. The IQ mutation simply increases repressor synthesis. Thus, the repressor accumulates more quickly and operon is repressed more quickly.
3
) 12 pts. One way that the spätzle (spz) gene product was identified as the Toll ligand in the Drosophila D/V patterning pathway, was by purifying active Spz protein and injecting it into the perivitelline space between the egg shell and the embryo.(a) Assuming that Spz protein remained at the site of injection, and that the injected embryos were derived from mothers mutant for spz, what would you expect the D/V phenotype of the injected embryos to be? (Answer for the embryo as a whole and for the site of injection.)
The embryos would be ventralized at the site of injection as the Spz protein would initiate local Toll signaling and thus establish a nuclear gradient of Dorsal protein whose highest point was at the site of injection. The rest of the embryo would be dorsalized as that's the phenotype in the absence of the spz gene since no signal is produced.
(b) Would you expect to find active Spz in easter- mutants? How about dorsal- mutants? Explain.
Easter is required to convert pro-Spz to the active ligand. Thus there should be no active Spz in easter mutants. Active Spz should be produced in dorsal mutants since the processing event is upstream and independent of dorsal function.
4) 38 pts. In E. coli utilization of ß-glucosides such as salicin requires three genes encoded in a single operon: bglG, bglF, and bglB. The bglB gene encodes an enzyme needed to metablize ß-glucosides--it has no regulatory role.
The bgl operon is normally expressed at very low levels, but it can be induced by ß-glucosides, all of which act as true inducers. A transcription terminator is located between the bgl-promoter and the bglG gene. Under non-inducing conditions most transcripts stop prior to reaching bglG. Under inducing conditions most transcripts read through the terminator. The following model for bgl regulation has been developed:
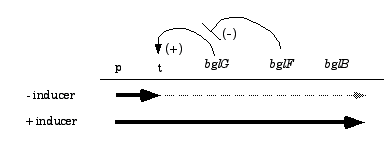
bglG encodes a transcription anti-terminator. The bglG protein is active in the presence of inducer and allows read through of the bgl "t" region.
bglF encodes a dual-function protein. It is a membrane protein that serves as the ß-glucoside transporter, and as a negative regulator of bgl operon expression. When inducing sugars are present bglF brings them into the cell, phosphorylating them as they enter the cytoplasm. In the absence of inducer, bglF phosphorylates the bglG protein, thereby inhibiting its transcription anti-termination activity.
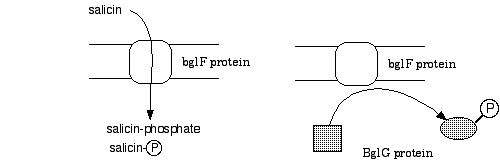
a) 10 pts. Fill in the following table stating the predicted Bgl phenotypes and whether the bgl operon would be induced, inducible, uninducible, or constitutive. Assume the mutations are recessive nulls.
|
Genotype |
inducer |
Bgl Phenotype |
||
|
absent |
present |
|||
|
wild-type |
Inducible |
Induced |
BGL+ |
|
|
bglG- |
Uninducible |
Uninducible |
BGL- |
|
|
bglF- |
Constitutive |
Constitutive |
BGL- |
|
|
bglB- |
Inducible |
Induced |
BGL- |
|
b) 8 pts. The model predicts that it should be possible to isolate additional types of bgl regulatory mutations. What sort of additional mutation(s) should you be able to isolate? State phenotype(s) and whether they would be dominant or recessive. Briefly explain.
bglG
mutations--non-phosphorylatable alleles. These should be constitutive as they can never be inactivated by blgF. They should also be dominant to bglG+.
bglF mutations--
alleles that cannot transport sugars but which can phosphorylate bglG protein. These should lead to a uninducible phenotype as bglF will always inactivate the bglG product. Such bglF alleles should be dominant as they will inactivate bglG even when a wild-type bglF product is present.
Alleles that can transport sugars but which cannot phosphorylate the bglG protein. These should lead to a constitutive Bgl+ phenotype. Such mutations should be recessive to bglF+ in diploid analysis. These are similar in regulatory function to bglF nulls, the only difference is that they can still use B-glucosides for growth.
One could also obtain mutations in the terminator. Theses should be cis-dominant constitutive alleles.
c) 8pts. Based on your answers to parts a) and b), suggest additional genetic tests of the model utilizing double mutants. Briefly state the predicted epistatic relationships and their meaning (i.e. what suppresses what). You do not need to consider all possible combinations--only present those that provide informative tests.
bglF- / bglG-
--the strain should be uninducible for bgl operon expression since the absence of the antiterminator will prevent transcription of the operon regardless of what happens with bglF, meaning that bglG- is epistatic to bglF-.bglG-constitutive / bglF-uninducible- --the strain should be constitutive for bgl operon expression since the non-phosphoryatable antiterminator will prevent transcription of the operon regardless of bglF's activity, bglG(con) is epistatic to bglF(unind).
The terminator mutations should lead to constitutive expression even when the uninducible bglG- and bglF(unind) alleles are present since there is no need for an active antiterminator when there is no functional termination site.
d) 12 pts. In vitro experiments were performed to test the model. Using a bgl+ template and pure BglG, BglF, and RNA polymerase holoenzyme proteins only, the following results were obtained 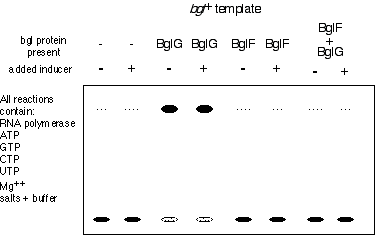 (as assayed by Northern blot.)
(as assayed by Northern blot.)
i) The results appear not to be entirely consistent with the proposed model for bgl regulation. Which data are consistent and which are problematic? Why?
All data are consistent with the model except those in the last lane. The problem is that BglF should not inactivate BglG in the presence of the inducer, yet it appears to do so here. The other data fit fine.
In the absence of the antiterminator protein trx stops at the terminator (lanes 1 and 2). In the presence of BglG, RNA polymerase reads through the terminator assisted by the BglG antitermination function (lanes 3 and 4.). (Note this should occur in the presence or absence of inducer as there BglG is active unless phosphorylated by BglF.) In the presence of bglF there should be no readthrough of the terminator as BglG is the antiterminator (lanes 5 and 6). With both BglG and BglF one would expect termination in the absence of inducer as BglF should inactivate BglG (lane 7), however, one would expect readthrough in the presence of inducer as BglG should be active (lane 8.)
ii) Can you think of a way to explain the in vitro results without discarding the model?
Propose a simple in vitro test of your explanation. (You may use any mutant protein or template and/or any "reasonable" biochemical reagent(s) you can imagine.)
The simplest explanation is that salicin must be transported through the membrane in order to inhibit the phosphorylation of the bglG gene product. Thus the inducer doesn't induce in vitro. One could test this by reconstituting the reaction in the presence of a membrane and testing to see if salicin induces read-through transcription in vitro.
Another test would be to use the BglF mutant that cannot phosphoryate BglG but which can transport salicin in the in vitro reaction. You would expect read through of the terminator under these conditions.
Extra credit 5 pts.
You have isolated an E. coli lac mutant with the following properties. The lac operon is highly expressed in the presence of IPTG or several naturally occurring B-galactosides but is off in the presence of lactose. This mutation is recessive in diploid analysis. Which gene is the best candidate to be mutated in this strain and what is the nature of the mutation?
The lacZ gene is the best choice. In order to convert lactose to the "true" inducer allolactose, it must be modified by the lacZ gene product B-galactosidase. Since the other inducers are also true inducers they require no modification. The lacZ mutant should be some sort of loss-of-function allele.
The lacY gene is also a reasonable choice. This answer, however, makes the assumption that all the inducers can get into the cell in the complete absence of the Y gene product. If that assumption holds then the operon could be induced by the true inducers but not by lactose, since lactose can never enter the cell.