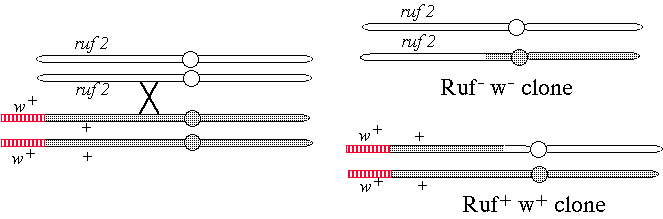
Note: Click on the figures to obtain a clearer image.
1) 12 pts. Consider the following pedigrees, all involving a single trait. Briefly explain for each case whether any of the following modes of inheritance can be excluded: X-linked dominant, X-linked recessive, autosomal dominant, autosomal recessive.
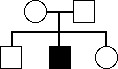
(a)
X-Dom: excluded, no affected parents.
X-rec: possible, mother could be heterozygous
A-Dom: excluded, no affected parents
A-rec: possible, both parents heterozygous.
(b)
(a)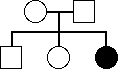
X-Dom: excluded, no affected parents.
X-rec: excluded, father would have had to be affected.
A-Dom: excluded, no affected parents
A-rec: possible, both parents heterozygous.
c)
X-Dom: excluded, all daughters should have expressed.
X-rec: possible, mother could be heterozygous
A-Dom: possible, male parent heterozygous.
A-rec: possible, female parent heterozygous.
2) 8 pts. Among native Americans, two types of cerumen (ear wax) are seen, dry and sticky. A lucky geneticist assigned to study the inheritance of this trait observed the offspring produced by different kinds of matings. The results were:
|
Parents |
Number of mating pairs |
Offspring |
|
|
sticky |
dry |
||
|
Sticky X sticky |
10 |
32 |
6 |
|
Sticky X dry |
8 |
21 |
9 |
|
Dry X dry |
12 |
0 |
42 |
(a) How is earwax type inherited? Briefly explain.
Earwax seems to be inherited as a simple dominant or recessive Mendelian trait. Sticky appears to be dominant and Dry is recessive. If so, crosses between "Dry" individuals should produce only dry offspring while crosses between sticky and dry and sticky and sticky would be expected to give rise to both types.
(b) Why are there no 3:1 or 1:1 ratios in the data?
In order to get Mendelian ratios you have to examine true breeding lines or individuals with known genotypes. In this case the Sticky individuals were unknown combinations of homozygous DD and heterozygous Dd individuals. Only the "Dry" d/d individuals can be expected to be true breeding.
3) 8 pts. A married couple went to see a genetic counselor because each had a sibling with cystic fibrosis. CF is a recessive disease and neither member of the couple nor any of their four parents are affected.
(a) What are the chances that their child will be affected with CF?
The change that either parent is a carrier is 2/3. Thus the chances of having a child with genotype cf/cf is 2/3 X 2/3 X 1/4 = 4/36 = 1/9.
(b) What is the probability that their child will be a carrier of the CF mutation?
The child would be a carrier if its genotype were Cf/cf (or cf/+).
This could occur if both parents were heterozygotes:
2/3 X 2/3 X 1/2 = 4/18 = 2/9
Or if one parent is heterozygous:
2/3 X 1/3 X 1/2 = 2/18 = 1/9
+
1/3 X 2/3 X 1/2 = 1/9
= 2/9
Thus the probability is 2/9 + 2/9 = 4/9.
4) 18 pts. Short Answer section: Answer the following questions with a short (one or two sentence) explanation or by the appropriate calculations.
(a) The red fox has 17 pairs of large long chromosomes. The arctic fox has 26 pairs of smaller shorter chromosomes.
(i) What do you expect to be the chromosome number in the hybrid somatic tissues?
Each parent would contribute its haploid complement of 17 or 26 chromosomes giving a total of 43 chromosomes per somatic cell.
(ii) What problems would the hybrid face in terms of mitosis and meiosis? Would you expect the animals to be viable and/or fertile? Briefly explain.
Because the homologs do not pair in mitosis, there should be no problem with segregation and the progeny should develop into adults.
Because many chromosomes would be expected to be unpaired they will not align in meiosis. In meiosis I the sister chromatids will both go to one pole leaving the other cell deficient for this information. With so many chromosomes it's vanishingly unlikely that any gamete will receive a balanced euploid set so the hybrids would be sterile.
(b) What is the standard used to establish linkage in LOD score analysis? What is the meaning of a negative (-) value for a LOD score?
A LOD score of 3.0 is taken as certain evidence of linkage. This indicates that the probability of a certain degree of linkage is at least 1000 X greater than the probability that the genes are unlinked. A negative LOD score means the assortment of the markers in the pedigree is better explained by independent assortment than by linkage.
(c) Recall the dominant piebald spotting trait in cats.
(i) When comparing the phenotype of one animal to another, what genetic term is used to explain the Piebald phenotype?
Piebald exhibits variable expressivity. Different S/s animals express different degrees of the white spotting phenotype, but all have some white spots.
(ii) From the perspective of comparing the individual hair-producing cells in an Ss cat, what genetic term would you use to describe the Piebald phenotype?
Individual S/s hair cells will be either white or colored. This is a qualitative difference and each cell has a variable penetrance of the phenotype. Some S/s cells express white others do not. For part (i) all cats express some white, but the degree of expression varies.
5) 12 pt. (a) Why do two different calico cats have their patches of orange and black fur in different places? Explain whether or not such a variegated coat pattern could occur in marsupials.
Note:
If you don't know the marsupial mechanism you didn't try very hard to solve problem 5 on the practice problems for the first exam. marsupial dosage compensation problem.Calico cats are heterozygous for the O and o alleles of the X-linked orange gene. Because of random X inactivation some cells express (O) Orange fur color and some cells (o) express black fur color. Because the inactivation process is random no two such cats look alike.
In marsupials, the paternally-derived X is always inactivated. This means that females will always express from the maternal chromosome so any individual animal would have a uniform coat color for an X-linked gene.
(b) In fruit flies, both eyes of females heterozygous for functional, partially functional, or non-functional white alleles are of uniform color. This is true regardless of the parental origin of the alleles. Does this exclude the common mammalian mechanism of X dosage compensation? How about the marsupial mechanism? If the mechanism is excluded, explain why; if not, explain and offer other evidence in favor of, or in opposition to, the proposed mechanisms.
The mammalian mechanism is excluded. Since white is cell autonomous, female flies would express a mixture of white and pigmented eyes if one of the X's were randomly inactivated.
The marsupial mechanism is possible given only the information in this problem. If females always inactivated the male X (and thus the white gene) they eyes would be a uniform eye color. However, from numerous examples in the text and lecture, you know that crosses between w/w females and +/Y males produce red eyed progeny which would exclude inactivation of the paternal X.
Extra credit--up to 4 pts. Two identical twin males marry identical twin women. What are the relationships between the couple's children if you're:
(a) a lawyer?
They are (first) cousins.
(b) a geneticist?
They are siblings.
(c) a headline writer for the Post?
I'll post your best ones after I grade the tests.
6) 18 pts. You wish to map the location of a new Drosophila X chromosome alteration called inv (for inviable) that was isolated after treatment with g-rays. You decide to map the lesion genetically. You set up the following preliminary cross:
![]()
The yellow gene (y) maps at the left end of the X chromosome at 0.0 cM and the Bar eye (B) gene near the right end at 57.0 cM. The Bar allele is a dominant mutation that changes the eye from its normal smooth oval shape to a bar shaped structure.
The cross produced the following male progeny:
|
phenotype |
number |
|
Yellow |
15 |
|
Yellow Bar |
50 |
|
Wild-type |
105 |
|
Bar |
330 |
(a) What can you say about inviable? Is there anything odd about its position? Show, or briefly explain, how you arrived at your answer.
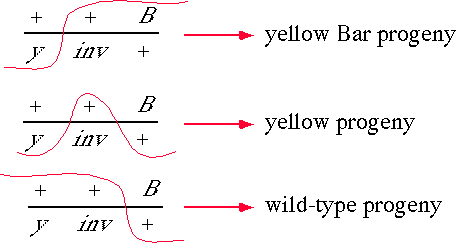
There were 15 + 50 y-inv recombinants out of 500 = 13% recombination
There were 105 + 15 inv-Bar recombinants out of 500 = 24 % recombination.
This would put inv 13 mu to the right of y at 13 cM and 24 mu to the left of Bar at
57-24 = 33 cM. Something is wrong if the lethal maps to two different locations.
The sum of the map distances between y and Bar in this experiment is 13 + 24 = 37 mu which is less than the expected 57 units. This implies that something was present that inhibited recombination. In theory, this could be a deletion or an inversion. Since the region is so large, 20 mu, a deletion would be lethal in the heterozygous state so inv is most likely an inversion with one breakpoint at ~ 13 cM and the other at ~ 33 cM.
(b) Predict the number and phenotypes of the female progeny.
If there were no inversion one would have expected independent assortment for genes this far apart. Since all females would have been viable you should have seen 250 yellow, 250 wt, 250 Bar and 250 yellow Bar females.
Since recombination is suppressed in the inverted region, you expect 37% recombination between y and Bar. Thus, you would predict 630 parental types and 370 recombinant types or 315 yellow, 315 Bar, 185 wild-type and 185 yellow Bar recombinants.
7) 24 pts. The insect compound eye is made up of a repeating pattern of hexagonal facets called ommatidia. A number of mutations can alter the structure of the ommatidia causing a disorganized eye phenotype that changes the normal smooth texture of the eye to a rough one.

Two new recessive mutations with identical "rough eye" phenotypes have been isolated. rufX maps on the X and ruf2 on the 2nd chromosome. The rufX and ruf2 mutations are thought to affect signaling molecules. One gene is predicted to produce an extracellular signaling ligand that instructs cells to adopt their proper eye fate. The other gene encodes a cell surface receptor thought to be required to sense the extracellular signal.
To test this hypothesis, the two ruf mutations were analyzed in somatic mosaics. For rufX, the X-linked white gene was used to mark the rufX mutant tissue.
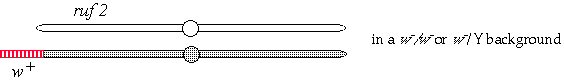
(a) Diagram how you would arrange the rufX and white alleles on the homologous chromosomes so that you could generate the rufX clones and identify them by their eye color phenotype. Explain your answer (a diagram may be most convenient.)![]()
Map of the X chromosome, white maps at 1.5 cM, rufX at 5.0 cM.
To generate somatic rufX clones that are marked with white- in a wild-type background the chromosomes should have been arranged as follows: w rufX / + +.
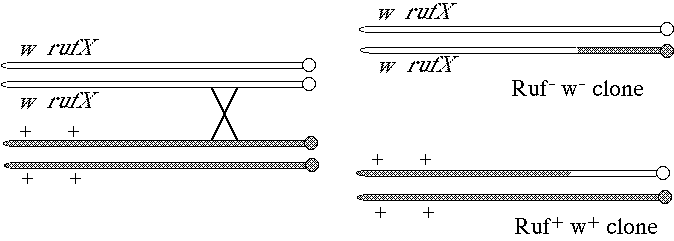
(b) The following figure shows the eye phenotype of a typical rufX mosaic. Is rufX likely to encode the extracellular signaling molecule or the receptor for that signal? Explain your answer.
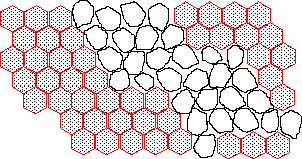
rufX most likely encodes the receptor molecule. All rufX mutant cells have the mutant phenotype suggesting the wild-type gene product acts cell autonomously. If rufX encoded the extracellular signaling molecule, mutant cells at the edge of the clone should have had the wild-type phenotype as their wild-type neighbors could have provided a functional signal.
This might have looked like:
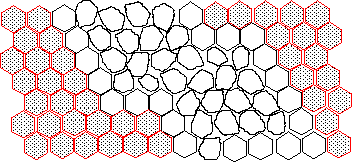
c) The ruf2 gene maps to the left arm of chromosome 2 where there are no eye color genes suitable for use in mosaic analysis. Can you think of a way that you could use white to mark ruf2 clones? You may explain with words or a diagram. Be sure to include all relevant genotypes. You may invent any reasonable genetic tool to assist you.
![]()