cape.
Why do chickens and cockatiels take up grit? To aid digestion. The
grit becomes lodged in the gizzard, which is a special chamber at the
rear end of the stomach. With its complement of grit, the muscular
contractions of the gizzard crush tough seed coatings (in the case of
chickens) and perhaps woody or cellulitic stems and branches (in the case
of dinosaurs) that have already been softened by
chemical digestion in the stomach. Thereafter, the food passes to the
intestine, the principal region of absorption and transfer of nutrition.
Seed-eating birds like chickens and cockatiels generally possess the
specialized chamber of the stomach called the gizzard, or more
technically, the ventriculus. This highly muscular organ is lined with a
hardened horny material called koilen. The lining makes the inside of the
gizzard somewhat rigid; the heavy muscles surrounding the koilen can
produce considerable pressure on the contents in the cavity of the
gizzard. This action crushes seeds and plant materials before they are
passed to the intestine for further digestion. Gizzards are found only in
seed-eating and plant-eating birds; meat-eaters and fruit-eaters do not
have (and do not need) gizzards.
Most, or perhaps all, birds with gizzards use grit to facilitate
digestion, although the function of the grit has never been fully
explained. Apparently, birds with gizzards take up grit when it is
available, but they can survive without it. Its real function is elusive.
Most biologists assume that grit somehow participates in crushing the
contents of the gizzard, a process called trituration by physiologists.
Exactly how the trituration of food in the gizzard is improved by grit is
an open question, however. Perhaps the grit rotates with muscular
contractions of the gizzard, slicing and cutting into the food.
Seed-eating birds have gizzards, and birds are descendants of
dinosaurs. Perhaps, then, plant-eating dinosaurs had gizzards. The
similarities invite comparison, but differences between birds and
sauropods suggest caution. For example, birds are toothless, but
sauropods had teeth. Modern birds with gizzards eat seeds, but few (if
any) Jurassic sauropods took seeds because nutritious seeds were not
available until angiosperms evolved in the Cretaceous Period. Sauropods
declined precipitously at the end of the Jurassic and survived only in
diminished diversity and number through the Cretaceous. The rise of the
flowering plants, the angiosperms, came at the very beginning of the
Cretaceous. Thereafter, flowering plants expanded dramatically; they are
now, by far, the dominant terrestrial plant.
Some paleontologists have suggested that Cretaceous sauropods, quite
unlike many other dinosaur groups, became increasingly restricted to relict habitats left over from the Jurassic.
Relicts of the Jurassic would be regions rich in ferns, cycads, and
conifers. There are modern analogues: for example, the rich fir and cedar
rain forests (with ferns blanketing the forest floor) that thrive along
the coasts of Washington and Oregon. Perhaps sauropods would do well
there. In any case, we do know that seeds of flowering plants were not
available to the sauropods of the Jurassic, when these plant-eating
dinosaurs reached their zenith.
There are other important differences between birds and dinosaurs, as
well. Birds are small and demand rich foods for their high metabolism.
Dinosaurs were large and, because their metabolic demands were lower,
probably did not require food as rich as did birds.
 |
| A herd of
Diplodocus. |
Overall, the bird analogy is useful but rough. Are there other
organisms alive today that use stones in their digestive tracts? Yes
there are: crocodiles, turtles, and some lizards. According to one study,
nearly all adult crocodiles of one population had stomach stones. The
function of stones in these carnivorous reptiles is debatable, at least
for crocodiles. Some authorities suggest that they are used as ballast
for buoyancy adjustments like the lead weights that scuba divers use to
make them sink. The digestive functions of stomach stones in crocodiles
may therefore be entirely incidental. But at least in crocodiles, the
stones are truly stones--not just small grit.
Similarly, the function of stomach stones may have been incidental in
dinosaurs, too. The hypothesis that stomach stones in sauropod dinosaurs
had no function (i.e., were only incidental) is as difficult to test as
the obverse, but it cannot be ruled out. Sauropod skeletons sometimes
contain stomach stones, but we cannot be certain that sauropods had
gizzards. Nevertheless, there is considerable circumstantial evidence
(especially from the excavation of Sam) that they had gizzards and used
stomach stones, the dinosaur equivalent of grit. Gastroliths
is a better term than stomach stones because the stones in
Sam's digestive tract may not have been confined to the stomach or a
chamber of the stomach like a gizzard.
Paleontologist Robert Bakker has argued for gizzards in sauropods and
for the use of stomach stones as an aid to digestion.
His conclusions, like those of many before him, are based on largely
circumstantial (and, in my view, weak) evidence, since no sauropod
skeleton before Sam has ever been fully documented as having gastroliths
in the visceral cavity.
Sam's gastroliths generally support Bakker's conclusions, but I am
still cautious in making physiological extensions from that evidence.
Nevertheless, Sam's excavation has furnished some exciting evidence that
can be used to bolster the arguments of gizzard advocates. It did confirm
that sauropods had (and presumably used) stomach stones, but this does
not automatically mean that they had a distinct chamber like a gizzard.
Absent soft-tissue evidence we cannot equate the function of dinosaur
stomach stones to the presumed function of such stones or grit in birds
and crocodiles. Indeed, I question the notion of any direct participation
in food grinding by stomach stones in the sauropod dinosaurs, which is
the function usually attributed to them. And the idea that they were used
for ballast is negated by the overwhelming evidence that sauropods lived
on dry land.
Sam had stomach stones--so many that they became a nuisance in the
excavation. We have identified and mapped more than 240, ranging in size
from about an inch to four inches in diameter, with a median size of
about two inches. Most were oblong, and some were spherical. A few were
flattened and roughly discoid. Each one was carefully exposed in the
sandstone surrounding the skeleton. Each was photographed, its position
plotted and mapped, and labeled before removal. Gastrolith documentation
was thus meticulous.
Some groups of gastroliths we excavated with the surrounding rock, to
keep them in position for later reference. Others were deliberately
removed in contact with bones, to preserve their original positions and
attitudes. Still others were unknowingly removed along with the bones in
the blocks we had established to ensure the integrity of the bones. Some
of these, surely, have not yet been "discovered" and won't be until the
bones are fully prepared for study with the surrounding sandstone
removed. With full preparation of the skeleton, we expect to find more, but the total will probably not be radically
different from the 240 we have logged to date.
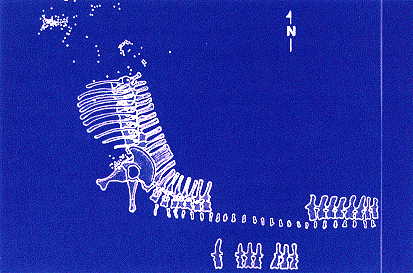 |
| Location of gastroliths. Simplified quarry map, with ribs shown
in original positions. Gastroliths were found mostly in two clusters: one
near the front of the pelvis, the other farther forward. In all, more
than 240 gastroliths were recovered, but not all are plotted on this
map. |
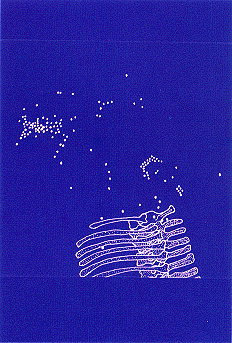 |
| Close-up of the anterior region of the skeleton. The semilunar
cluster of gastroliths at right center was protected from river currents
by a vertebra (not shown on this map so that gastroliths would be
evident) that had fallen on top of them. That bone ("Murphy's vertebra")
was dorsal vertebra no. 3. Also not shown here are the four isolated and
heavily eroded cervical vertebrae that were found (geologically)
downstream far to the right of this map. |
With the discovery of each stone, the progress of our excavation
slowed substantially; without these stones (or with a more casual
attitude as to their value) we could have completed the excavation of
Sam's skeleton at least a year earlier. But this unexpected bonus in the
excavation became a focus of attention when we realized that Sam's
gastroliths would be the first to be fully documented for any sauropod
dinosaur. And we knew that they could potentially play a role in
deciphering the burial history of the carcass, that they could assist
interpretation of Sam's anatomy, and that they might suggest something
about Sam's feeding habits--and that of sauropod dinosaurs in general.
How can we be sure that these stones are genuine gastroliths?
Skeptics, myself included, claim that most stones identified as
gastroliths are river rocks that were carried by streams and deposited
near the dinosaur skeletons with which they are associated. Therefore, by
this line of reasoning, gastroliths may not demonstrably be associated
with the dinosaurs. Their presence could be entirely coincidental,
perhaps owing to the carcass acting as a barrier to sediment transport or
perhaps owing to an eddying effect around the body, which would prompt an
abrupt drop in stream velocity, thereby causing stones sliding along the
bottom as bed load to drop in the sands surrounding the
skeleton. This abiotic explanation for the occurrence of alleged
gastroliths may be appropriate in many sites where dinosaur skeletons are
common. Thus, a demonstration that Sam's gastroliths are genuine stomach
stones, rather than impostors deposited by streams, cannot be marshalled
as evidence that all purported gastroliths are real. Instead, each
occurrence must be subjected to the same test: can we legitimately rule
out deposition by streams or other modes of deposition?
The gastroliths associated with Sam's skeleton surprised us. At first
we casually removed them from the excavation and mapped their occurrence.
Because no other sauropod excavation in my acquaintance had reported
gastroliths in direct association, I was unprepared to accept the first
cluster of stones that we found near the pelvis as an indication of more
to come. I was naturally skeptical then, but because Sam's skeleton was
buried entirely in sand, there seemed to be no other explanation.
Sam's skeleton lay in the middle of a layer of sandstone twenty feet
thick, bounded below by shale and capped above by a different, more
uniformly cemented and harder sandstone. Except for the gastroliths in
the pelvis of Sam's skeleton, no sedimentary materials larger than sand
were found in this ancient sand bar. If the stones, generally the size of
a plum, were stream-deposited, there should be gravels and cobbles
present in the sandstone, too, in gradational layers.
Ordinarily, stream deposits display gradations in grain size, a
feature geologists call "graded bedding." It is not unusual to find
gravels and cobbles associated with stream sands, but in vertical
sequence. Such changes in grain size are almost never abrupt; instead,
the sands are overlain by increasingly larger or smaller (clay-size)
sedimentary materials that reflect a change in stream velocity and
carrying capacity (bed load and suspended load). Where the change is
abrupt, without gradation, either no sediments of intermediate sizes were
available for transport (a rare and unusual situation), or other modes of
deposition must be considered: for example, rafting of cobbles caught in
floating blocks of ice or cobbles being carried to their resting place in
an animal's carcass. Deposition by stream action and
transport to the site in the body of a dinosaur are, however, the only
two credible explanations for the stones we excavated. Other
explanations, such as ice-rafting, lack evidence and need not be
considered as possible.
In our quarry stream deposition seemed a very unlikely explanation for
the stones. There were no gravel or cobble layers or "lenses" above or
below the skeleton, nor were there any within a radius of a hundred feet
from the excavation. The gastroliths in the pelvis lay directly on sand.
Under ordinary depositional circumstances, stream-deposited gravels and
cobbles dropped from a current would come to rest on other sediments
coarser than the sand. The fact that these stones were "matrix-supported"
was thus another important piece of evidence against their having been
stream-deposited.
All indications thus pointed to the conclusion that the stones had
been contained by the carcass, presumably in the digestive tract, that
they came to rest on the sand as Sam's carcass decayed, and that the
contents of the digestive tract spilled out and settled on the sand that
supported the skeleton. My skepticism was dwindling. I was becoming a
believer in sauropod gizzards. These were genuine gastroliths, presumably
from Sam's gizzard, and we were pleased to discover that we had real
documentation, for the first time, that sauropods (or at least Sam) did
indeed use stomach stones.
 |
| 101 gastroliths. These constitute a portion of the anterior
set, presumably associated with the crop. Many other gastroliths from
this set were collected in situ, with rock and bone, and could not be
included in this photograph. The largest gastrolith found in the entire
site has been placed in the center of this photograph, partially obscured
by another gastrolith. (Scale divisions are 1 centimeter.) |
Before the discovery of Sam's gastroliths only five sauropod skeletons were reported with associated gastroliths, and
the documentation in each case was meager. Three were in the Morrison
Formation in North America and were only casually described. Two were in
the Tendaguru beds of Tanzania, and the gastroliths were accurately
measured and described. In one of the Tanzanian excavations the
gastroliths were found in the neck region of a skeleton of Barosaurus. As
there were only five stones, these gastroliths were little more than
curiosities, but their position in the neck has an important bearing on
arguments presented below that Sam had a crop as well as a gizzard.
After this first discovery of a clump of two dozen stones in the
pelvic region of Sam, we did not expect to find more. The pelvis, where
this cluster occurred, was exactly where a gizzard would have been in
life--if the digestive tract of a sauropod resembled that of a chicken.
But a year later Peggy and Wilson Bechtel, the quarry excavation
supervisors, began to uncover more gastroliths--some in clusters--and by
the dozens. Like the ones found within the pelvis, these stones were all
matrix-supported, not associated in any way with gravels or cobbles that
would have been deposited by stream action.
With each new gastrolith the map of discoveries became more and more
tantalizing: the distribution of gastroliths around the skeleton did not
fit the notion that they were confined to a gizzard, and we were finding
many more than we expected. By the conclusion of the excavation in 1992,
we had mapped more than 240 gastroliths, distributed in two general
clusters: one tight group of about two dozen stones in the pelvic region,
and a larger, more scattered group farther forward, ranging from roughly
the middle of the rib cage toward the base of the neck. What did this
mean?
As I mentioned earlier, nowhere in the quarry or in nearby sediments
could we find any evidence to support an explanation that these stones
were river-deposited--or ice-rafted, for that matter. We therefore held a
working hypothesis that these were indeed gastroliths because they
couldn't be anything else. They were brought to this site in Sam's body,
where they spilled out as the carcass decayed. But this conclusion was
supported by several positive lines of evidence, as well.
First, we found sets of gastroliths, including one from the pelvis and
several from the more forward position, that came to rest in a line, in
contact with each other as though they had been contained by soft-tissues
when they came to rest on the sand. Some of the individual stones in
these sets were arranged in overlapping fashion, and some were on edge
rather than lying flat. These orientations are almost impossible in
stream beds, except where a stream bottom is mantled by river stones.
Deposition of single lines of stones in isolation seems to be nearly
impossible in stream-generated sediments.
This realization that the stones actually reflected the position of
stomach anatomy led to another surprise: some of these lines of
gastroliths lay well beyond the outline of the carcass as indicated by
the skeleton. They seem to indicate the position of entrails distributed
by a scavenger that had pulled the carcass apart for easier access. The
predator's tugging and pulling at the contents of the stomach brought the
gastroliths along too, and they were kept intact in folds of tissue that
lay on the bare sand. As the sand later filled in around the soft,
probably desiccated tissues, the gastroliths were held in their original
positions with respect to each other.
A second compelling piece of evidence supporting the stomach-stone
hypothesis was that some gastroliths were in direct contact with bone:
ribs and vertebrae. Rather than lying horizontally, some had an amazing
orientation. One stone, a flattened disk that I might have chosen as a
"skipper" to throw across the surface of a pond, was perfectly on edge,
in a vertical position. Others were similarly oriented. Stream action
could not have produced such startling positions for stream-deposited
stones.
 |
| The largest and smallest gastroliths. The largest gastrolith is
markedly bigger than all the others. Could it have caused Sam's death by
choking? (Scale divisions are 1 centimeter.) |
Third, measurements of these stones revealed another contraindication
of stream deposits. The statistical distribution of long-axis and
intermediate-axis dimensions of a natural population of river stones
deposited by a surge in stream velocity would be rather linear: many
small ones, fewer intermediate ones and fewer still of the very largest.
The sizes of Sam's stones, however, displayed a bell curve. A few were
small, the size of a peach pit. More were larger, with the highest
frequency being stones roughly the size of a plum. A few were somewhat
larger, the size of a small apple, and a very few were
larger still--the biggest being the size of a small grapefruit. Only a
very unusual stream could have deposited them.
We thus were left with only one reasonable conclusion: these stones
were indeed gastroliths, the indestructible dinosaur grit contained in
Sam's digestive tract at the time of death. This provides a suitable
standard for comparison with purported stomach stones from other sites,
even where the gastroliths are not in direct association with a skeleton
or their sedimentary context so easily discerned.
Satisfied that the sedimentary context is consistent with the
conclusion that the exotic stones associated with Sam's skeleton are
genuine gastroliths, we can deduce a great deal of information about
their function, their duration in the digestive tract, the anatomy of the
digestive tract, Sam's behavior in selecting suitable stones for
ingestion, and the history of the carcass between coming to rest on the
sand bar and its ultimate burial. (This last topic plays an important
role in the chapter on forensics. Here I will concentrate on
life-associated issues.)
 |
| Gastrolith texture. Several gastroliths here demonstrate the
extreme rounding and surface polishing that they undergo in the digestive
system. (Scale divisions are 1 centimeter.) |
 |
| More gastrolith texture. Here one of the gastroliths shows an
extreme development of rounding and polish. Its composition is
cryptocrystalline quartz. |
Sam's gastroliths are all rounded, and some are highly polished. Their
surface texture ranges from dull to waxy, in accordance with the degree
of polishing. Except for breakage during excavation,
none of the gastroliths have sharp edges; even the ones with irregular
shapes are highly rounded. Their color ranges from white to black, with a
rainbow of intermediate colors including shades of red, yellow, green,
and brown. Their mineral composition is less variable: all of the
gastroliths in this set originated as igneous or metamorphic rocks. All
of the gastroliths, moreover, are varieties of quartz (chert and
quartzite)--the hardest and most durable of the minerals commonly
occurring at the earth's surface. These facts are all important in
deciphering the function of the gastroliths and related aspects of Sam's
behavior.
The assumption that Sam acquired these stones from local river beds is
implicit here. That is the only reasonable source of exotic cobbles in
the Morrison habitat. These cobbles would have eroded out of rock
formations from localities more ancient than the Ojito site. Perhaps some
were repeatedly subjected to the sedimentary cycle of erosion and
deposition, with each rejuvenation of the landscape.
Sam most likely acquired these stones deliberately, while drinking at
a stream bank. On the other hand, it is possible that sauropods migrated
to source areas for particular rock compositions, an idea that I find
highly unlikely but which I cannot dismiss altogether. Several
paleontologists have suggested that identification of these source areas,
called their "provenance," might indicate the extent of migratory
movements among the dinosaurs.
According to some paleontologists, the quality of polish on a
gastrolith is a distinguishing feature that separates gastroliths from
stream-deposited stones. Therefore all gastroliths should have a high
polish, and ones that were in residence for a long time should be waxy.
The waxy trait seems to be a consequence of polish, on both the high and
low surfaces of the rock. Rocks with such a texture reflect more light
than rocks that have polish on the highs only, and they impart a striking
quality in reflected light. They feel waxy, too, in contrast to the
pitted and slightly roughened texture of rocks that lack this trait.
Not all of Sam's gastroliths, however, display a waxy surface. To the
unaided eye, they range from highly polished and waxy to dull and pitted.
The waxy ones are easy to distinguish from ordinary river stones, but the
dull gastroliths seem not to have lost the surface texture from their
original condition when Sam picked them up as river stones in an ancient
stream bed.
The origin of this polish, especially the waxy texture, is difficult
to explain. If these stones were used for grinding food, as generally
claimed, then they should all be pitted and scratched as a consequence of
grinding against each other. If the polish came from chemicals in the
digestive tract--say, acids in the stomach--they should all have the same
degree of polish, or at least all rocks of similar mineral composition
should display similar textures.
Tumblers used by rock hounds to polish their stones are not a fair
analogue for a dinosaur stomach. Stones polished in a tumbler do not
polish each other; instead, the tumbler is furnished with a supply of
softer polishing materials (usually culminating with talc), that impart a
lustrous surface as the stones roll and tumble. Dinosaurs did not possess
such carefully managed polishing materials. I surmise that these stones
lay in folds and creases in the digestive tract, particularly in the
gizzard (and crop), and that their polish came from the gentle action of
muscular contractions.
I cannot explain why some are highly polished and others are dull.
There seems to be no correlation with mineral composition, size, degree
of rounding, or map position in the quarry. But I can guess, I suppose
that the rocks in the specialized chambers of the digestive tract were
ingested at different times, as with my cockatiels, which continually
replenish their supply of grit. If so, stones in longest residence would
be more polished than ones in residence for shorter times. Also, some may
have been subject to polishing action more intensely than others just
because of position in the digestive tract. One fold of the gizzard may
have been more active in its crushing function than another fold, so that
some stones in the gizzard were subjected to more of the polishing action
than others.
In scientific terms we might express these observations on degree of
polish as "a tendency toward high polish and eventually a waxy texture,"
but one must first exclude the possibility that the tendency is reversed:
a waxy texture originally at the time of ingestion; rough and pitted
after a long time in residence. Several experiments have documented the
high polish of gastroliths; the researchers conclude that the ones with
high polish, and especially the ones with waxy texture, are more polished
than river stones or beach stones.
Sam's gastroliths are all rounded, a condition more striking and more
universal than their polish. None are angular; none have sharp edges. In
technical descriptions these stones would be described as well rounded,
an extreme condition of rounding that occurs naturally in the rolling
motion of rocks and cobbles in stream or shoreline settings. The
mechanical action of the rolling and sliding of rocks
that are originally angular (say, cubic for discussion here) wears on the
corners and edges, gradually changing the shape from cubic to a cube with
rounded edges and corners. Eventually the original cube approaches a
spherical shape. Because of natural fractures in the original rocks from
which they are derived, rocks rounded by the tumbling and sliding in
rivers or on beaches tend to be more like squashed spheres than spheres
and discoid, like skippers or poker chips.
Of course in nature rocks never become perfectly spherical or even
flattened round by this process, but they approach that extreme: this
idea is expressed as "sphericity." The sphericity of Sam's gastroliths is
uniformly high. Compared with a selection of rounded stream gravels, even
those preselected for roundness, Sam's gastroliths are remarkable for
their rounding.
Were Sam's gastroliths rounded by rolling and sliding in the digestive
tract? Yes, but they were almost surely rounded to begin with, when Sam
picked them up, probably from a nearby stream bed. Then too, when feeling
the need for more stones, Sam likely exercised some degree of choice in
picking them up. In Sam's digestive tract, their sphericity increased as
a consequence of renewed mechanical abrasion. They were simultaneously
polished and rounded. Now, all these deductions seem reasonable and they
fit our expectations. But a pesky problem remains to be addressed: Why
are there no angular stones?
If Sam only occasionally and randomly picked up stones, then some
should have been broken and angular, reflecting the breakage and
angularity of stones in stream beds. In a sample of 240 gastroliths we
should expect to find a dozen or more angular stones, with sharp edges
not yet rounded by the action of the digestive tract, stones taken up
just before death that hadn't been subjected to mechanical breakdown long
enough to reduce the sharp corners and edges. None of Sam's gastroliths
have sharp, or even dull, edges or corners; all 240 are rounded. I take
this collective trait to indicate that all of these gastroliths were in
residence in the digestive tract for a long time, perhaps years. If so,
by my argument, they stayed in the digestive tract until they were almost
totally ground down, say to the size of peach pits, and passed through
the digestive system with food.
Even if Sam picked up river stones in quantity, we should see a
spectrum of rounding. A set collected eight months before Sam died should
have more rounding (and polish) than a set picked up only a month before
death; no such pattern of subsets of rounding and polish emerge from our
observations. More likely, Sam picked up river stones casually and
irregularly.
Considering that gastroliths are poorly documented, they have been
used disproportionately in deductive arguments concerning sauropod
behavior, feeding habits, and stratigraphy. All of these topics have a
bearing on our studies of Sam. For example, was Sam selective? Were some
river stones more attractive than others? Certainly a sauropod would
select a stone within some size range (too large and it couldn't be
swallowed; too small and it would pass through to no effect). Possibly a
sauropod would select stones of some minimum level of sphericity. These
choices could have been made by sight or by simply spitting out those
that felt wrong in the mouth. But some paleontologists have hypothesized
that sauropods selected for composition or color too--suggestions that,
in my view, move out of the realm of science and into fantasy.
Some popular accounts of sauropods include the suggestion that these
dinosaurs undertook long migrations to favorite collecting sites. There
is no evidence for this proposal. I see no way to distinguish between
river stones picked up in nearby drainages (but derived from sources
perhaps hundreds of miles away) and stones collected from afar and
carried long distances in the body cavity of a giant dinosaur. The
well-intended purpose in these claims is to test the idea that sauropods
engaged in long migrations; that may have been true, but gastroliths will
not settle the question.
Similarly, some have suggested that sauropods selected only
quartz-rich rocks for gastroliths. Again, I disagree. The fact that Sam's
gastroliths (the only sauropod gastroliths thoroughly documented) are all
quartz does not lead to that conclusion; instead, these may have been the
only stones to survive the vigorous action of the digestive tract.
Layered rocks would be easily disaggregated because of differential
susceptibility of individual layers to pressure and gastric juices; rocks
of other composition, such as limestones or shales,
would be easily broken down in the acidic environment of the digestive
tract. Such stones would not survive for long as gastroliths.
Some paleontologists have relied on moas for making projections of
sauropod feeding habits and the functions of gastroliths. Moas, the huge
flightless birds of New Zealand that became extinct owing to human
occupation of the islands, used gastroliths extensively, and they seem to
have been highly selective. These giant herbivores, weighing as much as a
half ton, may have selected only white stones; perhaps only white stones
were available or were the only available rocks sufficiently durable to
withstand the rigors of the digestive tract. Nevertheless, I question the
direct application of moa behavior to that of sauropods.
Similarly, my notions on residence time in the alimentary tract (the
stones remained until virtually ground down and destroyed) have a bearing
on interpretations of purported gastroliths in the Morrison and Cedar
Mountain formations that are not positioned directly in association with
skeletons. According to geologist Lee Stokes gastroliths occur in great
abundance in the Cedar Mountain Formation (lower Cretaceous) of the
western United States. This formation overlies the Morrison Formation in
much of Utah and Colorado, and the two are difficult to distinguish
except by subtle differences. One distinction, according to this
argument, is the general lack of gastroliths in the Morrison. I agree,
except that I am not convinced that the purported gastroliths are
properly identified.
Whatever their origin, the widespread occurrence of these
"gastroliths" in the Cedar Mountain Formation is problematic, because
sauropod dinosaurs (and all other dinosaurs) are rare in that formation,
whereas gastroliths are rare in the Morrison Formation, which contains
the world's greatest bounty of sauropods. Paleontologist Robert Bakker
has suggested that isolated gastroliths may be the only residual evidence
of the positions of skeletons long since dissolved or redeposited, an
idea I find attractive but hard to prove. If it can be shown that the
conditions for preservation of bone were less favorable in the Cedar
Mountain Formation, this idea would become more plausible and deserving
of further consideration.
Sam's gastroliths have been the subject of several presentations and
papers, but full description of all the gastroliths is probably years
away, as the ones still encased along with bone await laboratory
preparation and study.
To summarize the facts about Sam's gastroliths: they are all polished,
but not all to the same extent, and some are waxy. All are well rounded;
none have sharp edges. Some are markedly disk-shaped. All originated from
igneous or nonfoliated (unlayered) metamorphic rocks; none are
sedimentary or layered; and they range from white to black, including a
rainbow of colors. Three-fourths of the gastroliths are between one and
three inches at their longest dimension. Long/intermediate/ short axis
dimensions of the smallest are .83 inches x .67 inches x
.51 inches; and the largest is 3.74 inches x 3.58 inches x
2.87 inches. Large (greater than 3.10 inches) and small (less than 1.20
inches) gastroliths are uncommon.
Not included in the summary above are observations concerning the
positions of the gastroliths in the quarry. These facts are just as
important as the physical qualities of the objects themselves. Contrary
to expectations, Sam's gastroliths were not found in a single pile, as
though dumped from a bushel basket and immediately buried. Instead, Sam's
gastroliths were spread unevenly over more than 1,600 square feet. We may
have missed some in the excavation, especially in the early years before
we fully appreciated their implications, but the patterns of their
distribution seem clear. In Sam's carcass were two clusters of
gastroliths, one large set in the forward region of the digestive tract
and a smaller set in the rear region found within the pelvic bones. Only
the rear set, consisting of 26 tightly clustered stones, can be
attributed to a gizzard, the grinding chamber originating as a
specialized rear pocket of the stomach.
The set from the front of the body seems to be centered at the front
part of the chest cavity, near the base of the neck. Evidently the neck
and head were displaced after the gastroliths spilled out of the body
cavity, leaving the gastroliths to mark the temporary position of the
front part of the body. Between the front set and the tight cluster in
the pelvis was a barren region of at least four feet along the vertebral
column where no gastroliths were found. This separation
is some evidence of a specialized forward chamber in Sam's digestive
system, which I have called the crop in reference to similar anatomy in
grain-feeding birds such as chickens and cockatiels.
There are no discernible differences in roundness, polish, size, or
composition between the front set (from the crop) and the rear set (from
the gizzard). This fact is inconsistent with the usual function of the
crop as a storage chamber, where grains and plant matter collect in lumps
before passing to the stomach and gizzard for digestion. The
inconsistency lies not in the anatomical position, but in the fact that
the gastroliths are rounded and polished, and many are waxy. If the crop
were simply a storage chamber, then gastroliths should have no function
there. The fact that they are highly polished and rounded, like those
from the gizzard, indicates that the front gastroliths were equally
involved in processing food, a radical suggestion for the function of a
crop. One highly unusual bird, the primitive hoatzin of South America,
has a crop with a gizzard's function. Although the anatomy of one bird is
not definitive evidence that Sam had a crop, it does demonstrate that a
crop with a grinding function is possible.
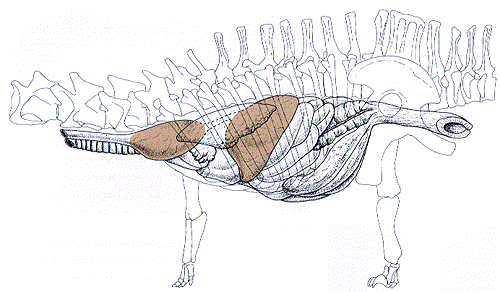 |
| The sauropod digestive tract. This schematic includes the crop
(the forward chamber at the base of the neck) and gizzard (between the
stomach and small intestine). |
Overall, we have sufficient evidence to propose that Sam's digestive
system resembled that of grain-feeding birds: esophagus, crop, stomach,
gizzard, and intestine. The crop and gizzard both contained gastroliths,
where food was probably pulverized in preparation for chemical digestion
(gastric secretion in the stomach and absorption in the intestine). Yet,
a nagging question remains: how exactly did Sam (and other sauropods) use
the gastroliths in digestion?
According to conventional explanations sauropod gastroliths were a
substitute for strong teeth. They were used in a grinding function in
place of chewing. I disagree, for several reasons.
First, not all sauropod skeletons seem to have had gastroliths, but
all sauropods had decidedly weak dentition. I doubt that stones as a
substitute for teeth were universally necessary for mastication. In
modern animals dentition is suited to diet. Only birds, which lack teeth,
have been documented to require grit as an accessory
material for grinding food--and only seed-eaters among the birds.
Second, gastroliths must have remained in the lowest creases and folds
in the capacious digestive organs, whereas food would have occupied the
chambers from bottom to top; only foodstuffs at the bottom of the crop or
gizzard would have been subjected to a grinding action of the stones.
Third, the bulk of the gastroliths is surprisingly out of proportion
to expectations: the 240 from Sam's carcass scarcely fill a ten-quart
bucket. To be important as an accessory grinding device, the bulk of
gastroliths should be considerably greater, or the foodstuffs must have
been forced through narrow constrictions blocked by gastroliths through
which the food had to pass before moving to the next chamber of the
digestive tract. For an animal weighing five or six times as much as an
elephant and processing many times as much food every day, the
gastroliths seem a weak addition to the digestive tract.
Instead, I propose that the gastroliths stirred digestive juices as
they rolled and tumbled in the bottoms of their respective chambers, much like a magnetic stirrer in a chemistry
laboratory. A small stirring device can create sufficient turbulence to
circulate fluids throughout a beaker, preventing unwanted settling and
segregation of materials by density. By this argument, gastroliths had a
similar role: to keep foodstuffs and the digestive juices mixed, not to
grind the food or pulverize it. I subscribe to this notion especially for
the crop, which was probably distensible and not highly muscular.
Fermentation and putrefaction in the crop might be controlled by adequate
mixing of digestive juices, preparing food for more vigorous chemical
treatment in the stomach. A grinding function for the gastroliths in the
gizzard seems more reasonable, where muscular contractions alone would
keep the food moving and prevent stagnation.
In both places, the rounding and polish of gastroliths would arise
from their contact with each other, the lining of the digestive tract,
and with gastric juices, not with the food materials; thus the grinding
(rounding and polishing) of gastroliths is a consequence of their
tumbling and rolling with each other, with only an inconsequential
grinding of food.
Interpretation of Sam's gastroliths has been a fascinating part of the
excavation of Sam. The gastroliths may tell us more about how Sam lived,
and whom among the living the great sauropods most resemble from a food
processing standpoint. But might the gastroliths also tell us how Sam
died?
I off-handedly remarked to a reporter that Sam's largest gastrolith,
the size of a grapefruit, was considerably larger than all the
others--and maybe it caused Sam's demise by lodging in the esophagus or
trachea. This tongue-in-cheek (or, stone-in-mouth) remark received
considerable publicity. Did Sam choke on this giant gastrolith? Maybe.
The evidence is far from conclusive, but the possibility cannot be ruled
out. There is, however, evidence suggesting another cause of death: Sam
may have been killed by a predator.
Seismosaurus