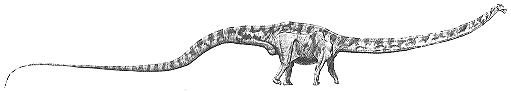
 7. Forensics with Sam |
7. Forensics with Sam |
Near Sam's carcass we found the tooth of a predatory dinosaur. Was Sam killed by a predator?
 |
| Tooth of a predatory dinosaur, found near Sam's carcass. This tooth is probably from Allosaurus or a close relative. It is the only indication of any other dinosaur at the site. |
In our judicial system, when there is suspicion of a wrongful death, specialists, usually including medical doctors, are called upon to reconstruct the crime: How did the victim die? What were the circumstances leading up to the time of death? What happened to the body after death? These are the questions asked by practitioners of forensic medicine. We shall do the same with Sam. As paleontological coroners, our mandate to determine the cause of death must be limited to the facts, including facts deduced from circumstantial evidence.
For paleontologists the court is our technical journals, where we submit articles (technically known as "papers") to present our case to other paleontologists. Truly, we are judged by a jury of peers. We become paleontological coroners because we want to know more than just the identity of the fossil organism. We want to know how it lived and how it died.
Once we piece together as much evidence as we can, we marshal our arguments in a logical, (usually) dispassionate fashion. Papers published later by other authors may cite us pro or con. This is a kind of forensic paleontology, and the principles are no different from those applied by medical specialists involved in criminology.
The technical name for forensic paleontology is taphonomy, and the specialists are taphonomists. The goal is to reconstruct the scene where the animal died as a historical account. Development of a plausible taphonomic history has been a persistent goal in paleontology almost since fossils were first recognized as evidence of past life.
In paleontology, as in forensic medicine, we are often faced with hundreds of facts or observations. We're never certain that we have collected all the facts or that we have chosen the right facts to record. Sometimes we overlook something that could be a critical link in unraveling a story. Such is often the case in mystery novels. Sherlock Holmes would have been an excellent paleontologist, an excellent taphonomist; in fact, his creator, Sir Arthur Conan Doyle, was an avid amateur paleontologist. Conan Doyle expressed considerable interest in dinosaurs and dinosaur tracks in The Lost World . He was even implicated in the Piltdown hoax (the excavation of a fraudulent skull and jaws of a hominid)--but that allegation was made without proof of complicity.
Exposure of the Piltdown bones as a hoax impressed on paleontologists the importance of acquiring new specimens only by excavating the fossils themselves--not by purchasing them from dealers or receiving them from a personal collection. Today's paleontologists, moreover, are equally concerned with the context (sediments and associated geological features), content (the fossil's anatomy and identity), and associations (other organisms). We are no longer satisfied with the simple recovery of a skeleton. The circumstances of its occurrence are just as important as the bones, and these are lost without meticulous attention to details that transcends interest solely in a skeleton. We must be detectives at the time of excavation, because the very act of excavation destroys evidence that can never be recovered.
In my supervision of the excavation of Sam I tried to impress upon the field crew the importance of being deliberate and thorough. I told the crew, "It's not a race." We collect as many facts as we can as we excavate, because by its very nature the removal of rock and sediment around Sam's bones destroys the clues that may be critical to understanding the life and death of this particular sauropod. For Sam we have lots of facts. Reassembling the evidence is something like solving a mystery, and in this case it could involve a killing. After all, it's possible that Sam was overcome by a predator, and we even have one tidbit of evidence to support that idea--the tooth of a predatory dinosaur.
First, let's assemble the facts and their immediate implications. All observations are appropriate, including what's not there. For example, there is no evidence whatsoever that Sam was an old individual or that death was due to old age: no injuries, no degeneration of bones from arthritis (which is known in some dinosaurs, but not Sam), no evidence that Sam was malnourished or diseased. None of Sam's bones were broken. There is no indication of steep topography, such as a cliff or a steep wall of rock, that might have caused accidental injury and subsequent death. The potential list of what's not there could be expanded for pages, but this partial list will suffice as a reminder that one must be aware of negative evidence in weighing the possibilities.
We know Sam's identity from our studies on taxonomy (see chapter 2): Sam was a large sauropod dinosaur related to Diplodocus and Apatosaurus, and Sam lived in the same area (what is now the Colorado Plateau) and same time (late Jurassic) as these two genera of herbivores. We do not, however, know Sam's gender. There are few clues among sauropod dinosaurs that call attention to sexual differences in the skeletal anatomy. Perhaps in the future we may be able to determine the sex of an individual from chemical evidence, but that's only a dream at present. If that technological advance should happen, however, the bones will be available for study--perhaps by a researcher in another museum or in a university. Curators regularly allow sampling for laboratory analysis much in the fashion of coroners exhuming a human body from a grave to make new tests for identity or cause of death.
Sam's vertebrae were lying on their right sides, and most of them were still articulated. Some of the ribs were still attached to the dorsal vertebrae, especially the ones on the right side, the lower ribs as the body lay in the ground. Other ribs, especially the ones on the left, or upper side, collapsed before the skeleton was buried. At least one of Sam's ribs was detached and carried a couple of yards away, perhaps by a strong current, but maybe it was flipped over by a predator (if there was one) or a scavenger.
Between caudal vertebrae numbers 1-7 (which were in articulation and continuous with the sacrum) and numbers 20-27 (the original eight excavated in 1985) was a gap in the line of the tail. A few fragmentary vertebrae, too eroded to provide much useful anatomical information, were found out of line but roughly in the position of the gap. We're not sure that we found all of the ones that belong in the resulting gap. At least five from that gap were there on-site when we began excavation in 1985. They were encased in heavily eroded sandstone mounds about six feet to the south of the main line of the east-west trending tail. Others seem to have eroded away entirely, so we will never have a complete set of Sam's tail vertebrae.
Accessory bones (called chevrons) in the tails of reptiles and kin, especially large dinosaurs, occur on the underside of each joint. In Sam's tail these chevron bones projected downward and rearward, protecting blood vessels and nerves underneath the tail and separating muscles on either side of the tail. The chevrons are considerably smaller than the vertebrae, so they could have been more easily moved by current action or by a scavenger.
Some of Sam's chevrons--at least three--were still in place when we excavated, that is, they were still attached at their respective joints in the tail, small bones next to large ones. This indicates that the bones were not separated by currents to any great extent, for if they had been carried by a stream, larger bones would have settled out first, smaller ones later, and the result would be a jumble of disconnected or even widely displaced bones. Actually, the hydrodynamic properties of bone are more complicated than simple distinction by size. Their shapes are also important: compact bones such as vertebrae are less easily moved by currents than long or flat bones such as the scapula or ribs. At least one other chevron, a large one from near the base of the tail, was stripped away from its original position and was found about a yard away. Like the displaced rib, this bone seemed to have been carried downstream slightly, as indicated by the slope of the bedding planes in the sandstone.
I once came upon a sea turtle skeleton on Padre Island near the southern tip of Texas, in the Gulf of Mexico. The bleached bones of the shell were stranded in the beach sands, still partially articulated, the dead turtle having been carried there by the high surf in a storm. The leg bones were nearby, just down current from the giant shell of the turtle. They had been carried away secondarily, after the turtle carcass disintegrated. I searched in vain for bones of the skull and feet. Being smaller than the shell and legs, these bones must have been light enough to be carried away by currents or wave action. Not realizing it at the time, I was a late-arriving witness to the taphonomy of this turtle's remains. They had been partially separated by wave action, after death and decomposition. If I had searched further, I might have found the skull and foot bones, but I had not yet learned to think critically about taphonomy. Moreover, I had come upon the skeleton as a beachcomber; it was not my professional quarry.
For Sam there are no leg bones or foot bones. As the excavation proceeded, we vainly searched to uncover some of the limb elements. It became clear, however, that the bones of the feet and legs were no longer associated with the torso or tail. Perhaps they remain hidden downstream in the Jurassic sand bar, beyond the limits of our quarry--just as my sea turtle leg bones were probably downstream from its shell. But if they are too far away, and even if found, they could not then be certified as parts of Sam.
The orientation and positions of the bones we did find can tell a great deal about Sam's death. As the bones were taken out of the quarry we plotted their positions on a grid map. Sometimes plotting the bones is difficult because we don't know exactly which bones are in a plaster jacket, and won't know until months or years later, so we plot the positions of the jacket on the map for future reference. Slowly the orientation of the skeleton takes shape on the map as the excavation develops.
Several important pieces of evidence about Sam's death can be seen on the map. First, the line of the vertebrae is curved in a tight arch. The curve is unnatural, as though the tail were lifted vertically. This is a rigor mortis posture, produced as the carcass lay on the surface of the ground without being pried apart by scavengers or buried too quickly beneath confining sands and muds.
Just after death, as an animal's body cools, the muscles and tendons stiffen, becoming almost rock hard. This stiffening is what causes the arching of the back. Muscles that hold the body up, acting against gravity in life, are larger than ones that pull the body downward. For this reason the muscles on top of the tail are more massive and therefore stronger than the muscles underneath. These muscles work in opposition to each other in life. The smaller ones underneath are assisted in their work by gravity, but after death with the carcass lying on its side, gravity doesn't help. The larger muscles on top "win" out in their stiffening after death. The rigidity is caused by the contraction of muscle fibers, the larger the muscles the greater their capacity for contraction.
The arching of Sam's back after death came about in this same way. The muscles above the tail and above the ribs and hips were stronger than lower muscles. The carcass remained unconfined by sediments, lying on the surface of the ground undisturbed. As the muscles stiffened the body arched backward.
One summer while excavating Sam's skeleton, we spotted a dead cow near a fence line on the range not far from the road. It had died only recently, for we had not seen the body the day before. Every day, as we passed "dead cow hill" I scrutinized the carcass. The hot summer sun caused the skin to desiccate and turn rigid. The legs became stiff and outstretched, and the body--originally collapsed--had gradually, over a period of several days, rotated to its side. The legs projected sideways, stiffened, and lifted from the ground.
By the fourth or fifth day, the body was not only stiff but quite bloated. The most peculiar change came with the head and neck. They had gone into the rigor mortis curve. Lying on its side, the head arched steeply backward, in a contorted posture. The heavy lifting muscles of the neck, situated on top of the neck bones, were more powerful than the lower muscles. The lifting muscles dominated in the rigor, creating this altogether unpleasant orientation of the carcass. Eventually the body fell apart. Vultures, crows, ravens, and other scavengers consumed the flesh, and the bones collapsed on the hard dry ground. They were never buried, however, so they never had a chance to become fossils. Instead they crumbled in the sun, and scavenging animals reduced them to powder and food.
Another important line of forensic evidence from the map of Sam's bones are the facts that the legs are missing, the neck is disconnected and most of it is missing, and the skull and jaws are missing. We were unable to determine where the legs are, but we do know that they are not connected to the body as they were in life. We found four neck bones downstream and disconnected from the main skeleton. They were heavily eroded from modern traffic (unknown to us, they were in the access road leading to the skeleton) and their positions cannot be determined with confidence. The neck bones we found were probably from the middle of the neck; the bones from the base of the neck, and from the head end were missing. Similarly, the bones of the head (the skull, jaws, and teeth) were missing, too.
Clearly, some of the skeleton had been separated before burial. Perhaps the missing bones are all downstream from the main body, buried in the sandstone only a few yards away like the rib, chevron, and four displaced neck bones that we did manage to find. The total area quarried is only about two thousand square feet. Every inch of that required hours of labor. We will always be haunted by the fear that just one more blow with the hammer-and-chisel on the sandstone might have exposed a missing bone.
Most of Sam's ribs are still connected at their joints on the vertebrae. The carcass was not entirely tugged apart by scavengers or torn apart by currents. Only one rib was disconnected, and it seems to have been flipped over once, coming to rest on top of the hip bones. The ribs of the upper (left) side all collapsed somewhat before burial, but the ribs on the lower side remained attached in their original positions. The rib cage had collapsed slightly before burial, but nearly maintained its original three-dimensional shape.
We have other important facts in this mystery. First, as the bones lay in the quarry, their upper surfaces (the parts facing toward the sky) are checked and cracked--not from our tools when we excavated, but from exposure after death. This deterioration is characteristic of bone that is exposed to air, like the cow's skeleton I watched deteriorate on the open range. Obviously some of Sam's bones were baked in the sun, cracking and flaking as they dried.
The bones were not completely desiccated, however. On the undersurface of one pelvis bone (the pubis), which stayed moist in contact with the ground, are several dozen tiny bore holes made by unidentified organisms, probably worms. They may have been feeding on organic material in the bone, such as proteins and fats. At least on that surface, hidden from the sun, there was enough moisture in the ground to support decomposer organisms.
The sediments, the sands around the bone, were all laid down by water. They were subjected to rather strong currents generated by a sizable stream or river. They resemble the sands deposited on the inner curves of streams as sand bars.
The texture and internal patterns of sandstones can provide geologists with a wealth of information concerning stream direction and velocity at the time of deposition, and whether the current was persistent or short-lived, giving way to gentler currents or even quiet water. In slowly moving water, suspended particles smaller than sand settle out and fall to the bottom as a layer of silt, because the turbulence is not strong enough to keep them moving. Such deposits become siltstones. In quiet water, even microscopic clay particles can settle out of suspension, leaving a muddy surface at the bottom. These may become mudstones or shales. Around Sam's bones there was no evidence of silt or clay deposition--only sand. Stream currents in the river where Sam's carcass was buried were thus strong and persistent, with little fluctuation.
That is the evidence derived from what is left of Sam. But was anyone else at the scene? This brings us to the stray tooth of a carnivorous dinosaur, perhaps Allosaurus or a close relative. It was found three feet downstream from the sacrum. The tooth is fairly large, from a predator that weighed at least a couple of tons and resembled the formidable Tyrannosaurus.
This tooth shows that a predator was there and that Sam's carcass attracted at least one meat-eating dinosaur. The tooth could not have drifted in on a current; otherwise, similarly sized rocks and pebbles would be scattered in the sandstone too.
Dinosaurs, like modern reptiles, grew continually through their life span. Their jaws grew continually, too, and with them grew teeth. They did not have permanent teeth, like those of adult mammals; instead, their teeth never fused to the jawbones. New teeth grew beneath the functional exposed teeth. As an old tooth wore out, or loosened in a feeding session, the attachment in the socket weakened and the tooth would fall out. New teeth growing in the jawbones soon replaced the missing teeth. Teeth of predatory dinosaurs are often found near carcasses of their prey, a permanent record of one dinosaur's feast.
I learned about this aspect of reptile anatomy in a rather memorable way as a senior biology major at Michigan State University. On a herpetology field trip to a marshy area in the northern part of the state, Professor Hensley demonstrated with a blue racer how to subdue an excited, thrashing snake caught by its tail. An hour later I glimpsed a rapidly disappearing tail in the wet bog. I lunged, knowing it was a harmless but excitable water snake. As though prompted by our professor, it began thrashing uncontrollably, but I had a strong grip near the end of the tail. I slung it up in a broad arc, and brought it down rapidly, the centrifugal force causing this irritated female (laden with eggs) to stiffen. As instructed, my arc carried her head downward and between my legs. I closed my legs together, pressing her long slender body between my thighs as I pulled her forward in one quick motion in order to grab her head.
My execution was perfect up to this point, but I was too slow. As I pulled her head between my legs, she reached sideways and took a powerful bite on the inner side of my thigh. I was startled, for I had naively expected her to cooperate, but I was not defeated. She finally gave up the struggle as I grasped her bulging midsection and neck with a firm grip. We identified her and released her back to the marsh. Later, I found several teeth lodged in my thigh, and several others in my levis. I had learned first hand about the manner in which reptiles shed their teeth.
Now, back to Sam: from the evidence at hand, can we assert that the meat-eater killed our giant herbivore? No, not with certainty, because it is likely that meat-eating dinosaurs did not always kill what they ate: if they found a carcass with flesh, they surely took advantage of a fast and easy meal. So, on the basis of the tooth, we arrive at two alternatives on the cause of death: Sam was either killed by a predator or Sam died of other causes and was then scavenged by a carnivorous dinosaur. I think the second possibility is most likely. I have a hard time imagining how a two-ton predator could kill a prey animal ten or twenty times heavier. Young or weak members of the species, yes; but the magnificent Sam, no. The predator may, of course, have been acting in a group, like a pack of wolves. This idea has been presented recently by dinosaur artists, but with little real evidence that predatory dinosaurs cooperated in the hunt. Even so, the scale of Sam to an Allosaurus would have been similar to that of a lion and an elephant; even prides of lions do not attack a healthy, adult elephant.
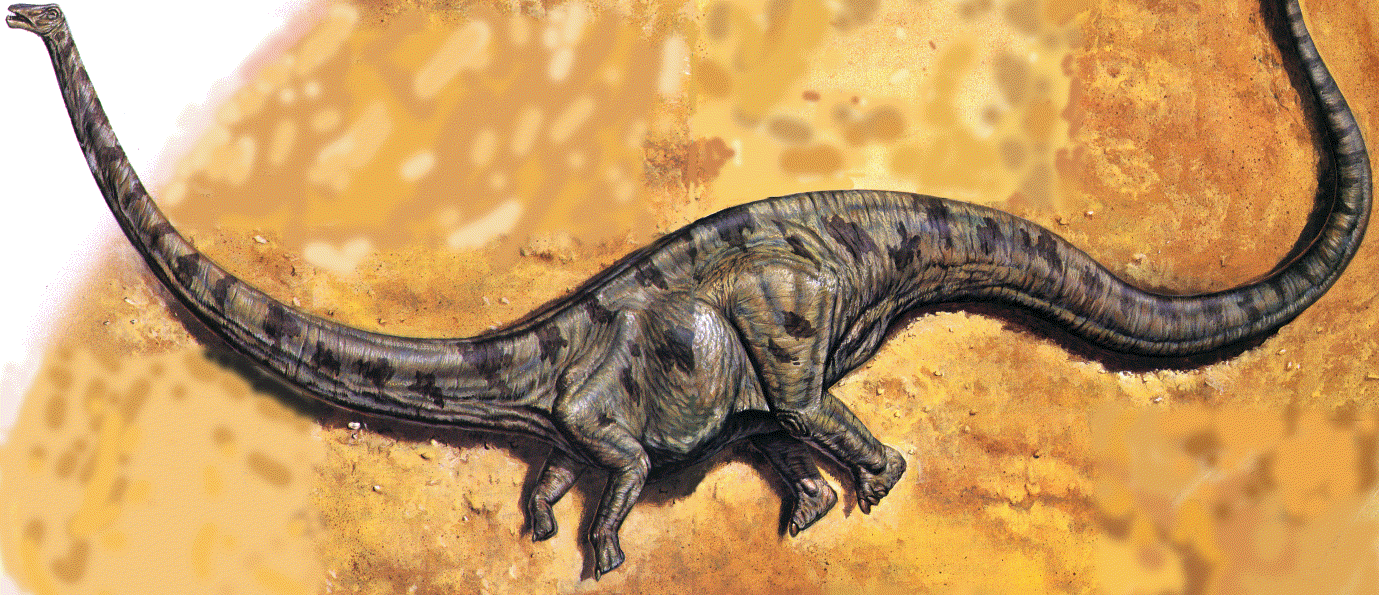 |
| Taphonomic history of Sam's body, from death to burial. A: death, carcass floating in shallow floodwater. |
 |
| B: stranded on a sand bar, with rigor mortis setting in; arching of neck and tail caused by stiffening of muscles as carcass dries and becomes rigid. |
 |
| C: scavengers, crocodiles and a dinosaur (possibly an Allosaurus), tugging the carcass apart, spilling internal organs and with them the gastroliths (the presence of crocodiles is conjectural). |
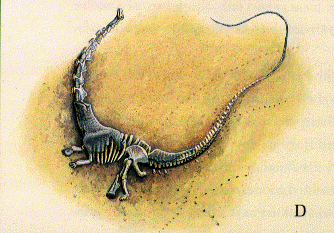 |
| D: disintegration of the carcass, leaving bare skeleton, some bones retaining tough connective tissue that holds joints in place. |
 |
| E: rejuvenated stream current displaces some bones of the skeleton. |
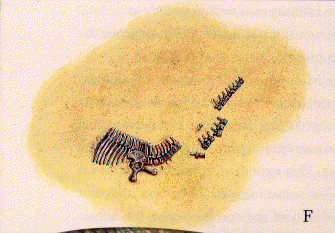 |
| F: skeleton is finally buried beneath a thick layer of sand and lies entombed for a hundred and fifty million years. |
At the very least, the tooth indicates that a meat-eating dinosaur was there and lost a tooth on site, just as the racer's teeth in my thigh indicated a snake had been there. The presence of the snake's tooth did not mean the snake had tried to eat me--nor, for that matter, I it. Likewise, the carnivorous dinosaur that owned that tooth until the time it fell out of its socket may not have been interested in feeding at all; the association of its tooth with Sam's carcass could be coincidental. What evidence might we accept as proof that a meat-eating dinosaur fed on Sam's flesh? Gouges in Sam's bones, or a tooth embedded in a bone might be clinchers, but such evidence is rare and we have none this convincing for Sam.
But, assuming for the moment that the presence of the tooth is evidence of a real association of a carnivore (whether acting as predator or scavenger) with Sam, one very useful induction can be made: the portion of the skeleton we discovered had not moved since the time it had flesh. That is, the skeleton was not secondarily carried away by currents after the flesh had deteriorated. For this reason, we suspect that the missing parts of Sam's skeleton could be found nearby. Even after we declare the excavation finished, which means that we give up the search for missing bones, we or other paleontologists will continue to watch the site for new bones to appear as a result of erosional processes, the same processes that initially exposed the original eight tail bones. In this way, perhaps decades from now, more of Sam's bones may be recovered.
The distribution of the gastroliths, described in the previous chapter, documents the same conclusion: the carcass came to rest here, its flesh was stripped away, and the stomach stones spilled out of the digestive tract. The bones of the rib cage and the back were not moved again; otherwise, the gastroliths and bones would not be found together. The scatter of the gastroliths indicate that a scavenger tore apart the visceral cavity, then tugged and pulled at its contents, eviscerating the carcass. With the evisceration came the gastroliths, redistributed by the actions of the scavenger as the guts were consumed or rotted. This pattern in turn leads to another speculation: the scatter seems extraordinary for a single scavenger acting alone. More likely, several scavengers competed for position on the carcass, and one or several may have tugged the entrails and pulled them apart at a safe distance from the main carcass. The skull and neck may have met a similar fate.
The gastroliths are important in our investigation for another reason: Sam may have choked to death, as mentioned in the previous chapter. The giant gastrolith crudely approximates a sphere in shape, like a contorted grapefruit. Even with alignment of its long axis parallel to the throat in swallowing, the bore required for the safe passage of this stone through the esophagus to the crop would have been prodigious in comparison with the bore necessary for safe passage of the next largest and all the others. The giant gastrolith is anomalous. Albeit an unorthodox idea, this scenario merits serious consideration.
Now, let's piece together these facts and their possible implications to reconstruct Sam's last days as a live animal, and the first days of the carcass in its long journey to becoming a fossil.
With all this evidence, we still cannot identify a cause of death. As discussed above, Sam may have been killed by a predator, or death may have come by choking on the largest stomach stone. Both ideas are plausible and can't be eliminated, but neither is satisfactory. I doubt that I could convince a jury that one or the other is the actual cause of death. Sam may well have died for some other reason.
Our forensic uncertainty is not unusual: in only a few cases in the fossil record can we identify exactly how or why an animal died. But we can reconstruct the history of the body from the time of death until it was buried.
The site of death could not have been far away. Sam may have died right where we found the skeleton. Alternatively, perhaps Sam died on the banks of a river with a fast current that picked up the carcass and carried it to our site, where it became stranded on the sand bar. This explanation might account for the missing legs, which for some inexplicable reason became separated from the remainder of the carcass before it was buried. We have no useful information concerning the fate of the leg bones, making this idea attractive even if it's not easily supported.
The current must have subsided abruptly, leaving the carcass stranded above water level on the sand bank, oriented so that the tail pointed downstream, its torso and neck taking the brunt of the current. Before deterioration of the carcass had gone far, it lay exposed to hot sun and dry air. The muscles arched the back into a rigor mortis curve as the skeleton lay on its right side on the sand bar. Tendons and muscles were still on the skeleton at this point, and the gastroliths were still in the belly.
Soon, perhaps within a day or two, one or more large predatory dinosaurs, such as Allosaurus, discovered the carcass and scavenged a big meal. Here was a windfall for a carnivore, enough meat to last for days or weeks. As the scavenging continued, Sam's stomach bloated, spilling its contents, including the gastroliths, onto the sand bar. The scavengers loosened the joints in their quest for meat, especially the joints of smaller bones and the upper ribs from the left side of the rib cage, now dangling in the air.
Upper surfaces of the bones were stripped of their flesh, and they lay exposed to the hot summer sun. The bones dried and began to shrink. Their surfaces bleached and brittle, the bones began to crack and flake in the heat.
A storm rejuvenated the meandering, slow-moving stream, which was flowing east toward modern-day Texas. As the water level rose, sand began to pack in and around the slowly submerging skeleton. The bones still had dried muscle and tendon, holding the joints together. Ripples and sheets of sand covered the gastroliths at base level and surrounded the bottoms of the bones. The partially decomposed carcass blocked the stream flow; eddies began to swirl around the edges of the skeleton as water was diverted by this unusual barrier. Steep cross-bedding developed on the lee side of the skeleton, coalescing with other cross-bedded layers in this persistent sandbar.
The turbulent stream undercut some of the bones, especially the tail. Sections that had lost their connecting muscle settled in the sand, gently slipping away from the main line of the tail. Other parts of the tail, still held together by tendons, remained in place on their sides, in the rigor mortis curve. Leg bones, if they were attached at the time the carcass came to rest at this point, were undercut by the current action, too, and they were rolled downstream, away from the vertebrae and ribs. The neck became detached at the same time, perhaps sliding downstream a few yards until sand buried the long line of slender neck bones, maybe with the skull still attached. One vertebra near the base of the neck fell forward and downward on its face, onto a pile of gastroliths that had spilled out of the belly earlier.
Layer upon layer of sand piled around and on the bones, now under water. Where the bones were cracked and flaked, sand worked into crevices. Loose bones, such as the chevron and rib, tumbled end for end until they came to rest on the gentle downstream slope of the sandbar. They, too, were covered by the accumulating sand. Soon the entire skeleton was entombed, buried until one hundred fifty million years later, when Arthur Loy saw the bones on the edge of the mesa in the Ojito Wilderness Study Area of New Mexico.
This is the best story of the death and burial of Sam that we can construct for now. But laboratory preparation and study of the bone blocks themselves has only just begun, so the story may change--or be restated later with greater confidence.
Although taphonomy as applied by modern paleontologists usually embraces the period only from death to burial, events long after burial are also part of the history of Sam's bones. And so other questions arise: what happened to the skeleton after it was buried? How, specifically, did it become fossilized? Is any of this bone original, or is it all "replaced"?
The answers are surprising.
 |
| Sam's demise. The carcass of an adult Seismosaurus, initially dried and dehydrated, now submerged by floodwaters in a summer storm. An opportunistic Allosaurus tears into the newly softened flesh. This scene is a plausible dramatization of Sam's preburial history. The crocodiles are conjectural. |