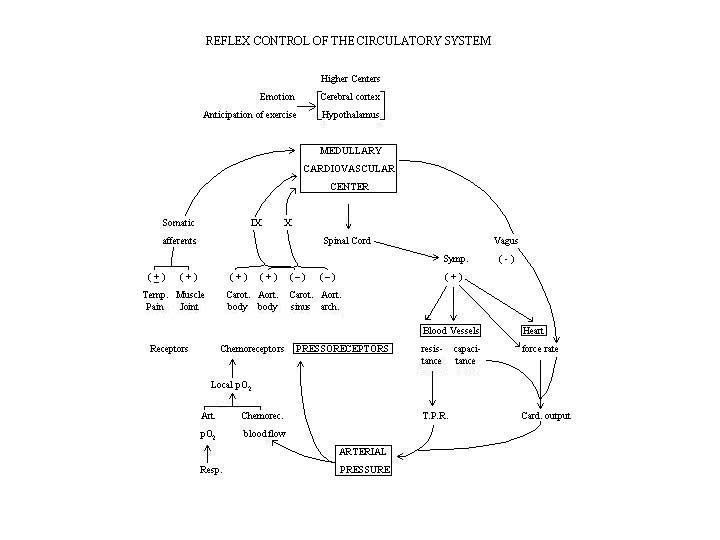
The above discussions on factors regulating cardiac output and peripheral resistance indicate that autonomic nervous activity, especially that of the sympathetic system, exerts important influences on cardiovascular function. These neural control mechanisms are discussed below in terms of the three major components of a reflex arc: the efferent system, the integration center, and the afferent system.
4.1 THE EFFERENT SYSTEMThe efferent control of the cardiovascular system is exerted through the autonomic nerves. The main parasympathetic influence on the cardiovascular system is a slowing of the heart rate by vagal impulses. The sympathetic adrenergic system, on the other hand, has a much wider variety of actions. These include the acceleration of heart rate, increase of cardiac contractility, reduction of venous capacitance and constriction of resistance vessels. The first three effects cause an increase in cardiac output, whereas the last action raises the peripheral resistance. Therefore, sympathetic adrenergic impulses cause an elevation of arterial pressure, which causes an increase in blood flow through regions (the brain and the heart) not constricted by the adrenergic influence.
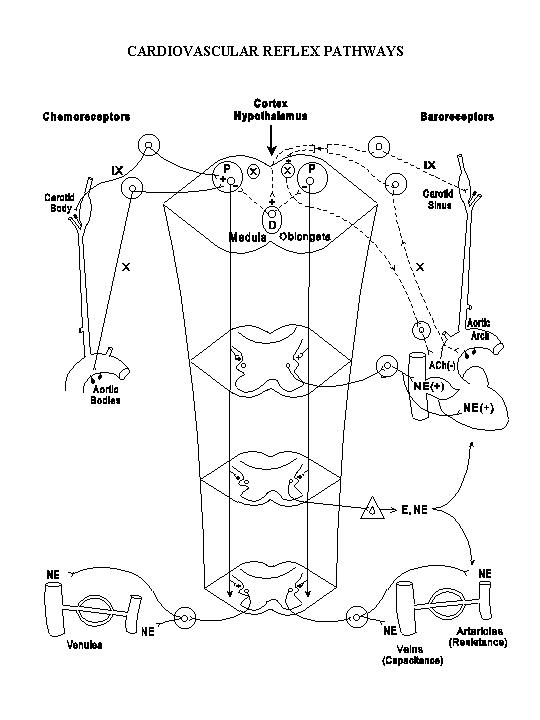
Circulatory reflexes are integrated at various levels of the central nervous system. A certain degree of integration of sympathetic reflexes can be achieved in the spinal cord. For example, sensory impulses from thermal or pain receptors in the skin can directly excite the sympathetic neurons in the lateral horn of the thoracolumbar cord. The most important level of integration of autonomic efferent activities to the cardiovascular system, howevcr, resides in the medulla oblongata. The cell bodies for the vagal efferents are located in the dorsal motor nuclei of the vagus in the dorsal aspects of the medulla. The neurons controlling the spinal sympathetic efferents are situated in the dorsolateral reticular formation of the medulla (see the following diagram). Electrical stimulation of these areas causes an enhancement of sympathetic adrenergic activity and an elevation of arterial pressure. Therefore, these bilateral areas are designated as the pressor areas. More medially and ventrally located is the depressor area. Stimulation of neurons in the depressor area causes an inhibition of sympathetic adrenergic activity and a decrease in arterial pressure. There exists a reciprocal relationship between the pressor area and the depressor area in response to afferent impulses, such that whenever one is activated the other is reciprocally inhibited. The pressor area and the dorsal motor nuclei of the vagus together constitute the cardiovascualr center. When we speak of an excitation of the cardiovascular center, however, we usually mean an excitation of the pressor area, while the other two areas may be reciprocally inhibited. The pressor and depressor areas send descending fibers through multisynaptic pathways to regulate the activity of sympathetic neurons located in the thoracolumbar cord.
In addition to the medulla oblongata, there are also neurons in the higher parts of the central nervous system which can modify sympathetic influences on the cardiovascular system. These include the neurons in the cerebral cortex (especially the limbic system) and the hypothalmus. They are particularly important in the activation of sympathetic cholinergic efferents in response to emotional excitement and anticipation of exercise. The hypothalamus not only contains neurons which can modify sympathetic adrenergic efferents via their influence on the medullary cardiovascular center, but also has neurons which send fibers directly to the sympathetic cholinergic neurons in the spinal cord. This hypothalamus-sympathetic cholinergic pathway plays a significant role in inducing sudden vasodilation in the skeletal muscle in response to certain stimuli (e.g., that occurring during fainting at the sight of blood in some individuals).
There are many receptors which send afferent fibers to the cardiovascular center. Some of these receptors reside within the cardiovascular system, whereas others are located outside of the circulation.
A. THE ARTERIAL BARORECEPTORS
In the systemic arterial system there are two sets of receptors which respond to changes in arterial pressure level. One set is the carotid baroreceptors located in the wall of the carotid sinus, which is situated at the origin of the internal carotid artery at the carotid bifurcation. The other set is the aortic baroreceptors located in the wall of the arch of the aorta (see diagram on cardiovascular reflex pathway). The afferent impulses from the carotid sinus travel in the glossopharyngeal (IXth cranial) nerve, whereas those from the aortic arch are conveyed by the vagus (Xth cranial) nerve. The baroreceptors are stretch receptors which are activated when the vessel wall is stretched by the arterial pressure. These receptors are not only sensitive to the absolute pressure level, but also to the rate of change of arterial pressure. Thus, the frequency of afferent impulses increases when the pressure is rising during systole and decreases when the pressure is falling during diastole (see diagram below).
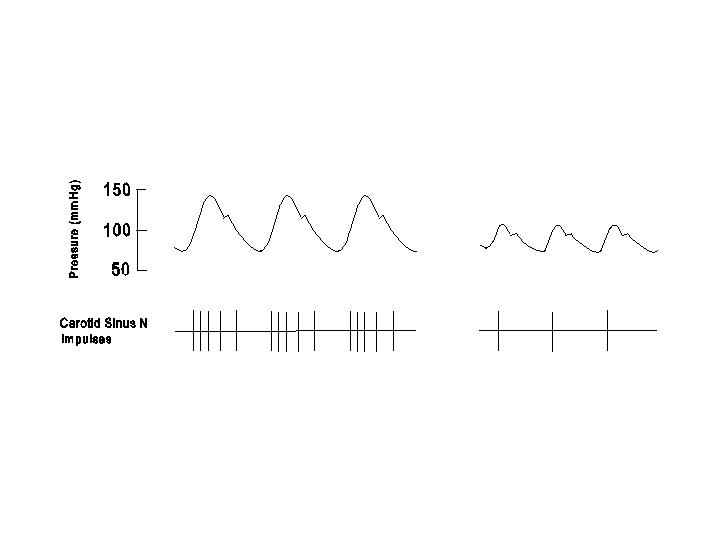
When the carotid sinus is perfused at various levels of steady pressures (no pulsation), the impulse frequency shows an S-shaped relation to the pressure level (see diagram below). The baroreceptor impulses cease completely at pressure below 50 mmHg, rise progressively between 60 and 180 mmHg, and reach a maximum frequency at pressures higher than approximately 180 mmHg.
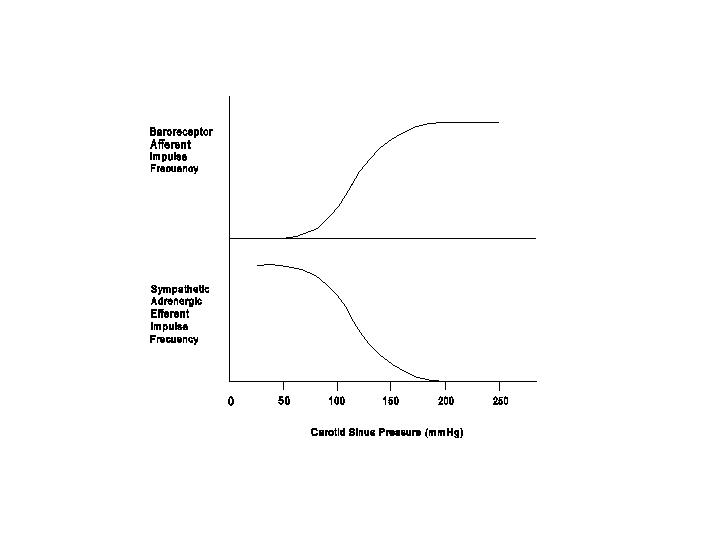
A very important feature of the arterial baroreceptor impulses is that they are inhibitory to the cardiovascular center (i.e., inhibition of the pressor area and excitation of the depressor area and the dorsal motor nuclei of the vagus). The normal arterial pressure of 120/80 mmHg causes a low-level stimulation of the baroreceptors,which send in inhibitory impulses to keep the sympathetic adrenergic impulses partially in check. A decrease of arterial pressure, e.g., due to blood loss, would reduce the inhibitory baroreceptor impulses and hence increase sympathetic adrenergic activity, thus tending to raise the arterial pressure. An increase in arterial pressure, e.g., due to cold induced vasoconstriction, would increase the inhibitory baroreceptor impulses and hence reduce sympathetic adrenergic activity, thus tending to lower the arterial pressure.
Therefore, the baroreceptor reflexes provide a negative feedback system to buffer any significant changes in arterial pressure. These reflexes are very important in maintaining the arterial pressure within narrow limits on a short term basis. Because of the tendency of the baroreceptors to adapt to a static level of arterial pressure, they have lesser importance in long term regulation of arterial pressure.
B. THE ARTERIAL CHEMORECEPTORS
The arterial chemoreceptors are located in areas very close to the baroreceptors: The carotid bodies are situated at the carotid bifurcation, and the aortic bodies are located near the aortic arch. These chemoreceptors are connected to the main arterial tree by arterial branches which carry a very high rate of blood flow considering the small size of the chemoreceptors. The afferent impulses from the carotid bodies travel in the glossopharyngeal (IXth cranial) nerve and those from the aortic bodies are conveyed by the vagus (Xth cranial) nerve.
The chemoreceptors are sensitive to their local metabolic environment when altered. Thus, these receptors are stimulated by decreases in local pO2 and pH and an increase in local pCO2. Because of the existence of a very high rate of blood flow per unit weight of chemoreceptors normally (approximately 2,000 ml/min/100 gm), these local metabolic factors are sensitive primarily to variations in the corresponding parameters in the arterial blood. Thus, a decrease in arterial pO2, an increase in arterial pCO2, or a reduction in arterial pH, which are changes occurring after suppression of respiration, may cause stimulation of the chemoreceptors. The chemoreceptors can also be activated when there is a severe reduction in chemoreceptor blood flow, e.g., following a large volume of blood loss.
The primary effect of chemoreceptor impulses is to stimulate the respiratory center and increase ventilation, but they also cause excitation of the cardiovascular center. The increase in sympathetic adrenergic activity resulting from chemoreceptor stimulation is mainly exerted on the resistance vessels to cause vasoconstriction.
When circulatory and respiratory functions are normal, the existence of a high chemoreceptor blood flow and normal values for blood gases and pH provides a local metabolic environment which gives rise to the normal chemoreceptor firing rate. When the arterial pO2 is reduced due to hypoventilation, the chemoreceptors become increasingly activated. The resulting chemoreceptor reflex causes a stimulation of respiratory movements to improve arterial pO2 and a selective constriction of resistance vessels to provide the brain and the heart with sufficient blood flow and oxygen delivery.
C. OTHER CARDIOVASCULAR RECEPTORS
(1) Mechanoreceptors.
There are receptors located in the atria and ventricles which are sensitive to mechanical stretch. The afferent fibers travel in the vagus nerves and in the sympathetic nerves.
There are several types of atrial receptors. The type that has the clearest physiological function is the receptors which fire upon atrial distention. An expansion in blood volume would cause atrial distention. The impulses from these atrial receptors inhibit the medullary cardiovascular center to cause dilation of the resistance vessels and also inhibit the release of antidiuretic hormone by the hypothalamico-hypophyseal system. This latter effect would cause an increased urine flow, thus tending to reduce the blood volume toward normal. A reduction in blood volume would reduce the impulse discharge from these atrial receptors, thus causing sympathetic vasoconstriction and release of ADH. Atrial distention can also directly cause release of atrial peptide from the atrial myocytes. The atrial peptide would cause vasolidation and renal sodium excretion.
The ventricular mechanoreceptors are located in the ventricular myocardium. These impulses cause reflex bradycardia and vasolidation, leading to a decrease in arterial pressure.
(2) Chemoreceptors.
Injection of several drugs (e.g., veratrum alkaloids and nicotine) into the coronary arteries of the dog causes reflex bradycardia and hypotension, together with some degree of respiratory inhibition. This is referred to as the Bezold-Jarish reflex, the significance of which is not known in man.
D. RECEPTORS IN THE SOMATIC SYSTEM
In addition to the above receptors which are found in the cardiovascular system, there are also receptors in the skin, periosteum, skeletal muscles and joints which send afferent impulses to modify the activity of the cardiovascular center.
(1) Thermoreceptors.
Upon exposure to cold, the receptors in the skin send afferent impulses to stimulate the cardiovascular center and enhance sympathetic adrenergic impulses. Therefore, the arterial pressure rises in a cold environment. This can be demonstrated by the cold pressor test in which an individual immerses his forearm in a bucket of cold water (e.g., 10°C) for several minutes. The vasoconstriction in the immersed hand is due to a combination of the direct effect of low temperature on the vascular smooth muscle and the reflex activation of the sympathetic adrenergic system. The vasoconstriction in the contralateral, un-immersed hand and other parts of the body is due to the reflex action alone.
The afferent impulses resulting from exposure to warmth causes an inhibition of the cardiovascular center.
(2) Pain receptors.
Superficial, sharp pain, e.g., that inflicted by a cut in the skin, is associated with afferent impulses which stimulate the cardiovascular center and sympathetic adrenergic nerves.
Deep, dull pain, e.g., that elicited by scraping of priosteum by an orthopedic surgeon, is associated with afferent impulses which inhibit the cardiovascular center. Therefore, this type of deep, dull pain is often accompanied by a precipitous fall in arterial pressure.
(3) Proprioceptors in muscle and joints.
During muscular exercise, the rhythmic activation of propiroceptors in the exercising limb leads to afferent impulses which are excitatory to the cardiovascular center and the respiratory center.
E. CEREBRAL ISCHEMIC RESPONSE
The cardiovascular center is stimulated by local accumulation of CO2 and acid metabolites. This occurs when cerebral blood flow is reduced as the arterial pressure falls below 50 mmHg. The cerebral ischemic response involves intense sympathetic adrenergic discharge, leading to a marked rise of arterial pressure as a last line of attempt to defend against cerebral ischemia.
F. HIGHER CENTERS
The activity of the medullary cardiovascular center can be modified by descending impulses from the cerebral cortex and hypothalamus. Emotional stimuli can activate these higher centers, which in turn excite the medullary cardiovascular center, giving rise to increases in heart rate and arterial pressure. These higher centers are also responsible for the activation of sympathetic system that occurs in anticipation of exercise.
A diagram summarizing reflex control of the circulatory system is shown below.