6.1 ARCHITECTURE OF THE MICROVASCULAR NETWORK
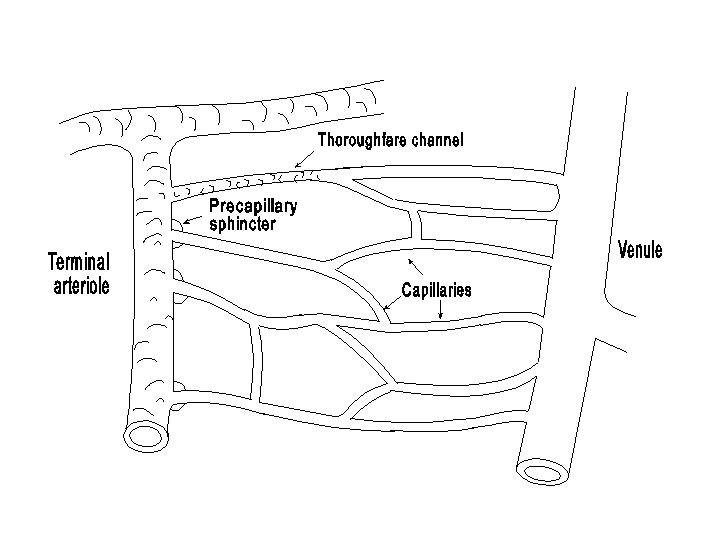
The vessels of the microcirculation consist of the arterioles, the capillaries and the venules. The arterioles, as mentioned before, are the major resistance vessels; the capillaries are exchange vessels responsible for transport of materials between blood and tissue; and the venules serve as collecting vessels, volume reservoirs and the site of postcapillary resistance. The detailed architecture of the microcirculation varies in different organs and tissues. An idealized diagram of the vessels in the microcirculation is shown in the following diagram.
The arterioles are approximately 50 microns in diameter with a single layer of smooth muscle, which provides the mechanism for control of vascular hindrance. The last part of the arterioles which feed into the capillaries are called terminal arterioles. The capillaries, which have a diameter of approximately 4-8 microns, possess only a single layer of endothelium and a basement membrane and are devoid of smooth muscle cells. In some tissues the entrance to the capillary is surrounded by a smooth muscle cell called the precapillary sphincter. The state of contraction of precapillary sphincters determines the number of capillaries receiving blood flow and participating in the transcapillary exchange process. In some vascular beds the connection between the arterioles and the venules is also made by some thoroughfare channels or A-V shunts, which are 10-15 microns in diameter and have smooth muscles covering their arteriolar end. Relaxation of these smooth muscles causes some shunting of blood flow through the thoroughfare channel away from the true capillaries. There are, however, also capillaries originating from the thoroughfare channel. On the venous side of the circulation, the capillaries coalesce into venules. A smooth muscle coat begins to appear in the medium sized venules, approximately 50 microns in diameter and larger.
An understanding of transcapillary exchange necessitates a rather detailed knowledge of the ultrastructure of the capillary. The capillary wall is composed of a single layer of endothelial cells. The lumenal surface of the endothelial cell is lined by a thin layer of glycoprotein (glycocalyx), and the tissue side is covered by the basement membrane. The roles of the glycoprotein coating and the basement membrane in controlling capillary permeability have not been established. The endothelial cell layer is probably the principal factor in the regulation of transcapillary movement, although the basement membrane may play a significant role in the renal glomeruli.
The adjacent endothelial cells in a capillary are separated by an intercellular space, the width of which shows regional differences. Depending on its morphological features, the capillary endothelium can be classified into three types (diagram below). The fenestrated endothelium has

regional thinning (fenestrations) of the capillary wall. Some of the fenestrations are completely open, whereas a thin diaphragm (closed) covers others. Fenestrated capillaries exist in the renal glomerulus and the intestinal mucosa. The continuous endothelium has no fenestration and the intercellular space contains a uniform spacing, with a width of 5-10 nm and tight junctions. The basement membrane is continuous. Capillaries with a continuous endothelium are found in the lungs, muscle and central nervous system. Discontinuous endothelium has wide intercellular spaces, approximately 50-100 nm in width, and a discontinuous basement membrane. Discontinuous endothelium exists in the spleen, liver and bone narrow. These structural differences in endothelium are associated with corresponding variations in capillary permeability (see below).
An important ultrastructural feature of the capillary endothelium is the presence of plasmalemmal vesicles. These vesicles, which are approximately 70 nm in diameter, are formed by invagination of the plasmalemmal membrane of the endothelial cell. Some of the vesicles are attached to the plasmalemmal membrane on both the blood side and the tissue side, whereas others appear to be free bodies in the cytoplasm but are in reality connected to the outside of the cell, (see following diagram). When the vesicle is attached to the plasmalemmal membrane, it has an opening of approximately 20-30 nm.

The transport of materials across capillary walls occurs mainly by two processes: diffusion and filtration. The driving potential for diffusion is the concentration gradient. The driving potential for filtration (or reabsorption) is the pressure gradient (fluid mechanical pressure and colloidal osmotic pressure). For a given potential, the rate of transport is a function of the characteristics of the membrane in relation to the properties of the substance being transported. Therefore, the ultrastructure of the capillary endothelium is an important determinant of transcapillary transport.
Diffusion across a membrane occurs in both directions. The net diffusion flux (Q), in moles per min, is given by the Fick�s law of diffusion:
where D is the diffusion coefficient of a substance through the membrane, A is the membrane area, ΔC is the concentration difference across the membrane (plasma vs. interstitial fluid) and x is the membrane thickness. The parameters on the right side of the equation are often grouped: (ΔC)/x is the concentration gradient, D/x is referred to as permeability and DA/x is the permeability-surface area product.
The endothelial cell membrane is lipid in nature, and substances with high lipid solubility, such as O2 and CO2 have high diffusion coefficients through the endothelium. This also means that the entire endothelial cell membrane area is available for diffusion. Therefore, a large diffusion flux occurs in response to a given concentration gradient.
Water soluble materials generally cannot diffuse through the membrane proper. Studies on the transcapillary transport of water soluble materials up to a molecular dimension of approximately 4 nm indicate that their diffusion takes place through aqueous channels (on the order of 10 nm) occupying a very small fraction of the endothelial surface (~0.1 percent in the skeletal muscle and ~1 percent in the kidney). This small area available for diffusion leads to a smaller flux of water soluble substances in comparison to lipid soluble materials.The intercellular junction is the ultrastructural basis for these small aqueous channels or pores.
The geometric relationship between the diffusing molecule and the channel is such that diffusion is severely restricted when the molecular dimension approaches 1/2 the channel size. Therefore, the fact that protein molecules larger than 5 nm can diffuse across the capillary wall indicates the presence of a large pore system. The ultrastructural basis of the large pore system is probably the plasmalemmal vesicles and the intercellular leaky junctions which are associated with endothelial cell turn-over processes. Because of the much higher protein concentration in the plasma, the large pore transport mechanism brings about a net transfer from the plasma to the interstitial fluid.
The above discussions apply primarily to capillaries with continuous endothelium. In capillaries with discontinuous endothelium or open-fenestrated endothelium, the wide intercellular space or open fenestration serves as the major large pore pathway for protein transport.
Physiological regulation of transcapillary diffusion is achieved mainly by controlling the available capillary surface area. In many tissues a significant fraction of the capillaries are unperfused in the normal condition. When there is an increased metabolic demand, the previously closed capillaries can be opened by relaxation of the terminal arterioles or the precapillary sphincters, a phenomenon referred to as recruitment
As formulated is Starling�s hypothesis, the fluid filtered across a capillary
membrane is proportional to the net filtration pressure which is a combination
of the fluid mechanical pressure (usually referred to as hydrostatic pressure,
but actually is the sum of hydrostatic and hydrodynamic pressures) and the
colloidal osmotic pressure. The rate of fluid filtration (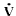 f in ml/min)
can be expressed as:
f in ml/min)
can be expressed as:
f = kf [(Pc - Pif) -
(πc - πif)]where kf is the filtration coefficient, Pc the capillary pressure, Pif the interstitial fluid pressure, πc the colloidal osmotic pressure in plasma, and πif is the colloidal osmotic pressure in interstitial fluid. Vf is positive for outward filtration from capillary to interstitial fluid and negative for inward reabsorption in the opposite direction.
The above equation is analogous to the Poiseuille-Hagen law, and kf is the reciprocal of filtration resistance, or the hydraulic conductance. Filtration occurs mainly through the intercellular junctions of the small pore system. Hence kf varies directly with the number of these junctions with the fourth power of the pore radius, and inversely with the pore length. It follows that kf is much higher in organs with discontinuous endothelium, e.g., the liver, than in structures with continuous endothelium, e.g., skeletal muscle.
The capillary pressure decreases continuously from the arteriolar end to the venular end.
The higher pressure in the arteriolar capillaries favors filtration, whereas the lower
pressure in the venular capillaries tends to cause fluid reabsorption. For the sake of
simplicity, we shall examine the mean capillary pressure (Pc), equivalent to the
pressure value somewhere in the middle of the capillary network, as a function of the arterial
pressure (Pa), venous pressure (Pv), precapillary resistance (Ra)
and postcapillary resistance (Rv). The blood flow () through a
region is given by:
| = |
Pa - Pc Ra |
Pc - Pv Rv |
Normally Pa-Pc is approximately 4 times the value of Pc-Pv in most tissues, hence Ra is also approximately 4 times the value of Rv. Solving for Pc from the two expressions of the right yields:
| Pa+Pv (Ra / Rv) 1 + (Ra / Rv) |
Therefore, the capillary pressure is a function of Pa, Pv and the ratio Ra/Rv. Pa and Pv are essentially the same for most areas in the systemic circulation, except in regions with several resistances in series, e.g., the mesenteric-portal hepatic circulation and the renal circulation (with afferent arteriolar, efferent arteriolar and venular resistances). Capillary pressure would rise with an increase in either arterial or venous pressure (if the ratio of arterial to venous resistance remains unchanged), but a given increment of change in venous pressure has a much greater influence than arterial pressure on capillary pressure, because the venous pressure change is magnified by the factor Ra/Rv (normally approximately = 4). Regional variations in Ra/Rv lead to corresponding differences in capillary pressure. In a given region the ratio Ra/Rv can be altered by neurohumoral and autoregulatory factors. The pressure in the capillaries will fall in response to a vasoconstriction upstream (↑Ra) or a vasodilation downstream (↓Rv). Therefore, an increase in Ra/Rv causes a decrease in Pc and a tendency for fluid reabsorption. On the other hand, vasodilation upstream (↓Ra) or vasoconstriction downstream (Rv), i.e., a decrease in Ra/Rv would cause an increase in Pc and a tendency for fluid filtration. This inverse relationship between Pc and Ra/Rv, which can be deduced by intuitive reasoning, can be made more apparent by rewriting the last equation as:
| Pa - Pv + Pv + Pv(Ra / Rv) 1 + (Ra / Rv) |
Pa-Pv 1 + (Ra / Rv) |
The shifting of Pc with variations in Ra and Rv is also shown in the following diagram.
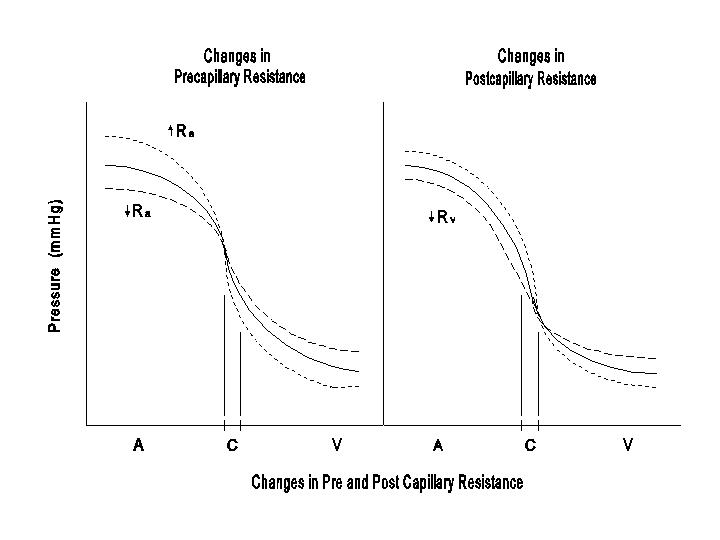
If these changes in Ra and Rv occur in a large part of the circulation, then Pa and Pv would change as illustrated in these diagrams. On the other hand, localized alterations in Ra and / or Rv would only affect the regional Pc without significant effects on Pa or Pv. The diagrams indicate that while changes in Ra and Rv affect the arterial pressure in the same way, they have opposite effects on capillary pressure and fluid exchange.
The interstitial fluid pressure (Pif) is much lower than the capillary pressure and varies in individual regions. It may even be slightly negative in some parts (e.g., the skin). Whether positive or negative, Pif is fairly close to zero, and the net fluid mechanical pressure (Pc - Pif) is dominated by Pc.
The colloidal osmotic pressures in capillary plasma (πc) and in the interstitial fluid (πif) are functions of the concentration and diffusivity of the proteins present. The relationship between colloidal osmotic pressure and protein concentration is not a linear one as described by the Van�t Hoff law, π = cRT, but is a nonlinear function of c:
where c is the molar concentration of the macromolecule, R is the gas constant and T is the absolute temperature, π rises increasingly steeply as the protein concentration is raised. The deviation of π from the Van't Hoff Law is partially due to the Donnan effect from macromolecular charges. The charges of the macromolecules immobilize counterions and these counterions are considered to be impermeable to the endothelium. This charge effect leads to the effective colloidal osmotic pressure to be greater than the magnitude caused by the non-ionized macromolecules, referred to as the Donnan effect. The plasma protein concentration is normally approximately 7 g/dl or about 1 mM, and the colloidal osmotic pressure is approximately 25 mmHg. The interstitial fluid protein concentration varies in different regions; it is higher in areas with discontinuous endothelium than in areas with continuous endothelium. In general, the interstitial fluid protein concentration is much lower than the plasma protein concentration, and the net colloidal osmotic pressure (πc - πif) is dominated by πc.
The actual colloidal osmotic pressure exerted by the proteins depends not only on their concentrations but also on the reflection coefficient (σ), which is a measure of the degree to which molecules are retained or reflected by the membrane. For small molecules which can diffuse freely through a membrane, none of the molecules are reflected; the reflection coefficient is zero and there is no osmotic pressure. For other molecules that show no permeability through a membrane, all of the molecules are reflected; the reflection coefficient is unity, and the osmotic pressure is fully exerted. For substances with partial permeability the reflection coefficient is between 0 and 1, and this coefficient indicates the degree to which the osmotic pressure should be discounted in the real system. Thus, the reflection coefficient is very high (close to 1) for large proteins in capillaries with continuous endothelium (e.g., skeletal muscle), whereas it is lower for small proteins (e.g., albumin) in capillaries with discontinuous endothelium (e.g., liver).
The above discussions indicate that the rate of fluid filtration should be written as:
The net effective colloidal osmotic pressure, σ(πc - πif), is low in the liver not only because of the high π if but also because of the low σ.
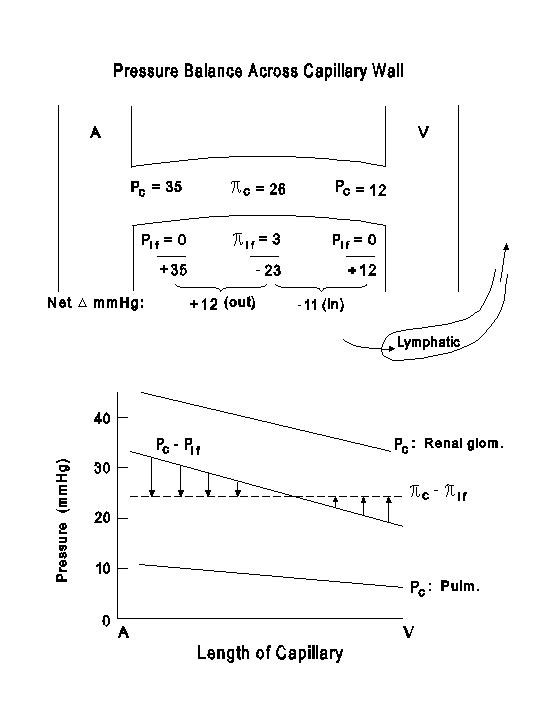
The net fluid mechanical pressure shows considerable regional differences. The liver has a low Pc because more than 2/3 of the inflow is via the portal vein, but the low net fluid mechanical pressure can effect a significant transcapillary filtration because of the existence of a low net colloidal osmotic pressure as discussed above. The renal glomerular capillaries have a high Pc because of the existence of the efferent arterioles downstream, and this favors glomerular filtration. The pulmonary circulation is a low pressure system, and the low Pc favors fluid reabsorption, thereby reducing the likelihood of pulmonary edema.
The next diagram illustrates the balance between fluid mechanical and colloidal osmotic pressures in the skeletal muscle capillaries. In the lower part of the diagram, the fluid mechanical pressures in the capillaries of the renal glomeruli and pulmonary circulation are also drawn for comparison.
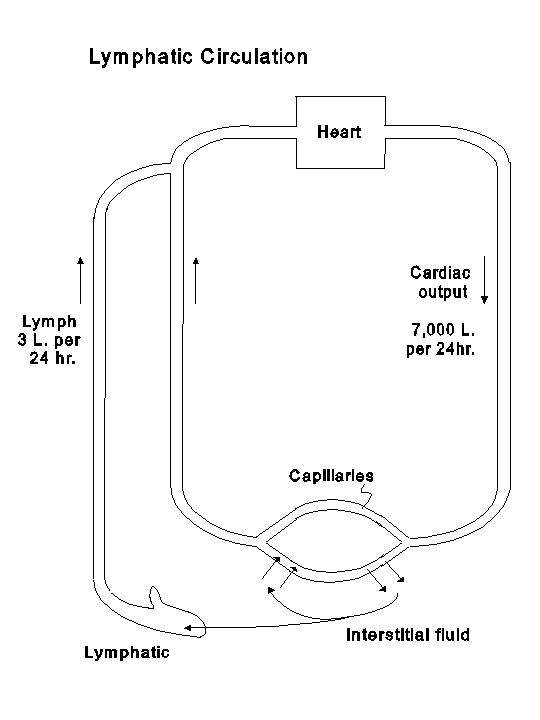
In transcapillary exchange in most tissues the outward filtration slightly exceeds the inward reabsorption, and the net outward filtration is normally drained away from the tissue by lymph flow. The lymphatic system begins as blind sacs in the interstitial space. The endothelium of lymphatic capillaries is discontinuous, and the basement membrane is not prominent and can even be absent altogether. Proteins, which have escaped from the blood capillaries into interstitial space, can enter the lymphatic capillaries. The larger lymphatic vessels do not have significant permeability for proteins. Therefore, the proteins are returned with water and other solutes via the lymphatic system (thoracic duct and other lymphatic ducts) into the large veins. The lymph protein concentration in different regions varies with the blood capillary permeability to proteins; it is high in regions with discontinuous endothelium and low in areas with continuous endothelium. The total lymph flow is probably over 3 liters per day, and approximately 100 gm of proteins pass through the lymphatic circulation per day to be returned to the systemic circulation. Therefore, as indicated in the following diagram, lymphatic circulation provides a circulatory path for fluids and solutes (including proteins) from capillaries through interstitium to the large veins.