The above experiments together make a convincing case that the basic organization of the simple cell receptive field -- its subfields, its preferred orientation, and some measure of its orientation selectivity -- are laid out by the spatial organization of the geniculate input. However, not all of the response properties of simple cells can be successfully predicted from this simple scheme.
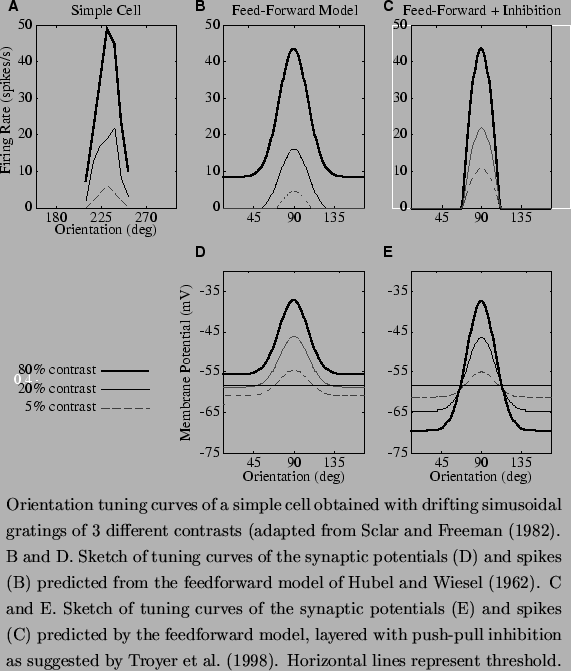
Perhaps the most serious inadequacy of the feedforward model is revealed by a striking feature of simple cell responses, the contrast-invariance of orientation tuning of the responses to drifting gratings (Skottun et al., 1987, Sclar and Freeman, 1982). As shown in Figure 4A, in real simple cells the width of the orientation tuning curve varies little as the contrast (strength) of the stimulus is varied. Only the height of the tuning curve increases with contrast. This behavior is difficult to explain in Hubel and Wiesel's simple feedforward model, because it is well known that the strengths of the responses of retinal ganglion cells (see Troy and Enroth-Cugell, 1993) and geniculate relay cells (Cheng et al., 1995) increase with the contrast of the stimulus. The difficulty is shown in Figure 4D, where the peak excitatory input that would arise from the geniculate input in response to drifting gratings is plotted as a function of orientation for different contrasts. A key feature of the model is that, even at non-preferred orientations, the depolarization at high contrast will be non-zero and will increase with contrast.
To understand this behavior, consider a grating at right angles to the cell's preferred orientation. Where it crosses an ON-region of the receptive field, the bright portion of the grating will activate the underlying ON-center geniculate relay cells at rates as high as 100 spikes per second. At the same time, the bright portion of the grating will suppress OFF-center inputs where it crosses the OFF region of the receptive field. But the reduction in the responses of the suppressed OFF-center cells saturates at zero spikes per second, and since the activity starts from a relatively low spontaneous rate of 10-15 spikes per second, the reduction cannot balance out the excitation of the ON-center cells. So stimuli of the non-preferred orientation above a certain contrast evoke a net excitation from the relay cells. Furthermore, the amplitude of these non-optimal relay cell responses grows with contrast, since the responses of the relay cells themselves grow with contrast. As a result (Figure 4D), the model predicts that the net excitation evoked by non-optimal orientations at high contrasts can exceed that evoked by the preferred orientation at low contrast.
Because both the baseline level of the predicted membrane potential tuning curve and the size of its peak grow with stimulus contrast, the feedforward model breaks down when an unvarying threshold is applied to derive the spike responses of the cell (Figure 4B). No matter where threshold is placed, the orientation tuning width of the resulting firing rate responses will show considerable broadening with increasing contrast, unlike the responses of real simple cells. This is the ``iceberg effect'' -- beneath the spiking threshold lies more broadly tuned excitatory input, which would be revealed by increasing contrast in the feedforward model. Furthermore, if the threshold is too high, stimuli at low contrasts will evoke no spike responses at all, whereas with low thresholds, stimuli at sufficiently high contrast will evoke responses at all orientations. Yet few simple cells show these behaviors (Figure 4A).
The contrast-invariance of orientation tuning has not been studied for stimuli other than drifting gratings, but similar problems exist for any stimulus. For example, consider a drifting bar. Even a low-contrast bar evokes spikes at the preferred orientation, while even a high-contrast bar does not evoke spikes at the orthogonal orientation. Yet the high-contrast bar will evoke net LGN excitation, because it will raise the firing rates of the LGN inputs it excites much more than it can lower the firing rates of the LGN inputs it suppresses. Since this net excitation increases with contrast, a sufficiently low-contrast bar of the preferred-orientation and a high-contrast orthogonally-oriented bar will yield the same peak level of LGN input, a level that must be suprathreshold given the spiking responses evoked by the preferred-orientation stimulus. Thus, the feedforward model would predict spiking responses at non-preferred orientations to high-contrast drifting bars.
In summary, if the signal underlying simple cell orientation tuning originates in feedforward LGN input, then to explain contrast-invariant tuning, some mechanism must raise the amount of LGN input required to yield a spiking response at higher stimulus contrasts.