BMEN E3500, fall 1998
These notes deal with transport and reactions that involve serial processes, either transport and reaction(s) in which all equations pertain to only one compartment or processes that occur through a series of compartments. Four reaction scenarios, which are interesting when at least some of them occur together, are:
No reaction. It is important to remember that the potential exists for many more reactions than one actually encounters in a given situation. A classic example is the expression of enzymes in cells that are catabolizing glucose to provide energy in the form of ATP. The reactions are nearly irreversible and thus inappropriate for glucose synthesis. The enzymes for glucose synthesis (gluconeogenesis) are not expressed. Likewise, when a cell is storing energy as glucose polymer (glycogen) the catabolic enzymes are not fully expressed and enzymes that implement a pathway that is different from the reverse of that used in glucose catabolism are expressed.
Irreversible reaction. It is a moot point whether irreversible reactions actually exist, maybe somehwat like the question of whether anything exists in just two dimensions. But just as things get pretty thin, reactions can show damn little reversibility: they lie so far to the right that one cannot detect reactant when they reach equilibrium.
Reversible reaction. There is a finite (not infinite, not zero) reaction in both directions.
Equilibrium. e.g. KAB alone, notkAB / kBA determines the relationship between product and reactant. For example, in the case considered below cB = KAB cA. A and B are "locked together" and a separate relationship for B is not needed.
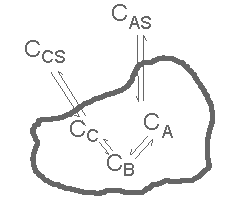
Here we analyze, in depth, a pretty simple case (that considered in class): See the figure above. Substance "A" exists in the surroundings of a compartment, permeates into the compartment, reacts to form "B" which reacts to form "C" which diffuses out of the compartment. The reactions are reversible, not a priori at equilibrium. To illustrate some basic points we don't put in additional compounds (D, E, F ...) but they could be there, allowing only F, say, to diffuse out. The intermediate compound B isn't allowed to diffuse out because a serial process is to be considered. If B were allowed both to diffuse and to react we would have parallel elements, easy enough to analyze but fogging up powerful conclusions to be drawn from results for purely series processes. The reactions are treated as first order, so that only linear equations need be solved. With a little computing power different eaction orders and Michaelis-Menten kinetics are still easy enough to analyze but the more complex results would, again, fog up some of the general conclusions to be drawn.
We apply the compartment conservation equation to each species, excepting AS and CS which are, respectively, treated as a source and a sink. We assume that A cannot react outside the cell and that AS and CS are not in equilibrium in the surroundings. Here asre the resulting equations:
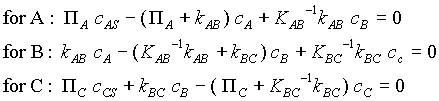
Considering the source and sink concentrations to be stipulated, there are 3 equations in 3 unknown concentrations. Notice that the reverse reaction rate constants have been written as the forward reaction rate constant divided by the equilibrium constant for the reaction. One can solve for the concentrations and then for the overall rate of reaction, N*:
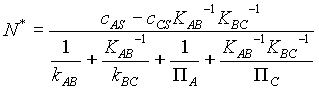
The quantities P i (where i equals A or C) are the membrane permeabilities multiplied by the diffusion or permeation area (which has been termed AD), divided by the compartment volume (which has been termed V). The point of this derivation is to study this equation. Notice that it can be considered as an example of the general form: flow equals driving force divided by a sum of resistances. The flow is N*, the rate at which A flows through the system.
(One could say as well the flow through the system is that of B or C, because at steady state and, as here, with a stoichiometric ratio of 1, the flows all equal each other.) It would be a good and fair test question to ask you how to correct these equations if one molecule of, say, A, produced two molecules of B.
The driving force is the difference between the concentration of A in the surroundings and the concentration of A that would exist if C were in equilibrium with A in the surroundings. Thus, if C were present at the concentration with which it would be in equilibrium with A, N* would be zero. Even though the reaction of A into B is possible because of the permeability of the cell and the enzymes that allow reaction to occur in the cell, no net reaction would, in fact, occur. The enzymes can only accelerate the approach to equilibrium; they cannot change equilibrium. If A and C are already at equilibrium in the surroundings, the cell cannot cause further reaction.
The resistance is (just as you saw it when you studied Kirchoff's laws in physics) the sum of the individual resistances, which are in turn the reciprocals of the "conductances", here the rate coefficients. Notice, however, that these resistances must be adjusted (using the equilibrium constants) for the fact that they are not all defined in terms of the same concentration (e.g. that of A), but rather are defined in terms of the different concentrations. These adjustments are not arbitrary; they come directly from solving the equations given above.
Below are the detailed solutions for the individual concentrations:
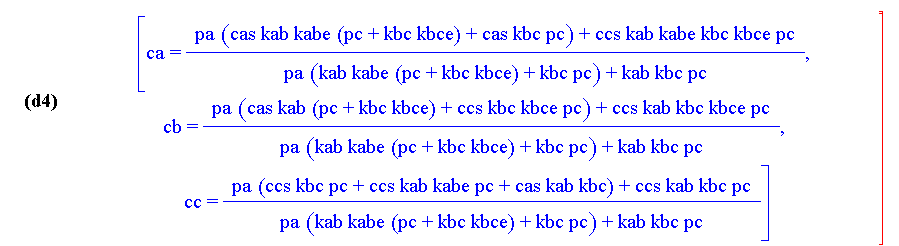
The result is from a computer algebra program, Macsyma, similar to Mathematica. The symbols have the following meaning:
| pa | cas | kab | kabe | pc | kbc | kbce | ccs | ca | cb | cc |
| P A | cAS | kAB | KAB-1 | P C | kBC | KBC-1 | cCS | cA | cB | cC |
You can use this information to solve for intermediate concentrations for this scheme. You should be able to derive similar schemes. Be sure to work on the related problem set.
Return to course main page.