
BMEN E3500, fall 1998
(assignment page scheduled for 11/7/98)
The compartment concept is founded on the idea that a finite, bounded volume in space
is under uniform conditions. Transport within a compartment ("intracompartmental
transport") occurs instantaneously and does not require any analysis. Transport among
compartments ("intercompartmental transport") occurs across all or part
of the boundary shared by two compartments. Here we consider in detail how to describe
transport among compartments. We divide this consideration into two parts: the topology of
intercompartmental connections and the physical-chemical mechanisms used to quantify
transport through these connections. One common and simple topology of intercompartmental
transport is that between one finite compartment and another infinite compartment that
completely surrounds the finite compartment. The second compartment is called, simply,
"the surroundings". Because of the infiniteness of the surrounding compartment,
it can influence, but cannot be influenced by, the finite compartment. Transport between a
single compartment and its surroundings is shown in Figure 1-A.
The directed lines may represent either convection or diffusion. Other topological arrangements of interest include two compartments, both finite, communicating only with each other (1-B). Compartments in series (1-C) comprise another frequent and important arrangement. Compartments may be arranged in parallel, (1-D), which introduces two other possibilities: unidirectional flow which, in practice, is most frequently realized when the dominant mode of transport is convection, and a through-flow: the compartments fed by a flow from a source and discharging to a sink. The source and the sink play the role of the surroundings in 1-A. Series-parallel arrays, 1-E, are also possible. Recirculating arrays (1-F) are frequently encountered, often in a compartmentalized system that uses a valuable constituent that is supplied or synthesized in limited quantity, and whose loss or leakage must correspondingly be limited. Combinations of these topologies are encountered. One very important combination, in which one finds the series, parallel, and recirculating topologies, is the compartmental representation of the human circulation, shown in Figure 1-G. (Only three of many parallel capillary beds in the systemic circulation are shown. Each of the compartments is considered to "communicate" with the others only by convection, which corresponds closely with actual observation.) Further consideration of how to analyze these topologies is deferred in order to consider, first, the physical-chemical mechanisms of transport. These will be developed and analyzed primarily in terms of one compartment communicating only with its surroundings.
Consistent with statements made in the introduction of the compartmental concept, the total area bounding a compartment can be divided into sub-areas that may allow: (1) no transport (are impermeable), or (2) convective transport to a degree that any concurrent diffusion through that sub-area is negligible, or (3) diffusive transport with either negligible or comparable convection.
Formal treatment of intercompartmental transport always involves only an algebraic expression. In the case of simple convection, the expression is simply the product of a flowrate between, say, compartment 1, the source and compartment 2, the sink, and the concentration in compartment 1, q12 c1. When significant diffusion is present, the algebraic expression relates the rate of transport to one or more coefficients (commonly called permeabilities), the area over which these coefficients act, and a set of conditions that exist within the participating compartments. A simple example: two compartments communicate through a shared area, A, and the transport of a substance I is governed by a permeability Pi and the difference in the concentration of I in the two compartments:
N*=NiA = PiA(ci,1 - ci,2)
This expression relates a flow, N*, to a coefficient, Pi, an area, A, and a "driving force", (ci,1 - ci,2). It is universal practice to describe all intercompartmental transport involving diffusion in the form of the preceding equations: a flow (describable either as the flow-proper, here, N*, or as the product of a flux here Ni and an associated area, here A) The flow equals the product of a permeability here Pi, the area, and a driving force, here, (ci,1 - ci,2)). Even very complex transport expressions, involving complications not yet considered here, are generally reorganized so that there is an easily recognizable permeability, area, and driving force. The reciprocal of the permeability-area product is sometimes used and is called a (transport) resistance.
The following discussion of transport mechanisms is concerned with two matters:
1. The effect on transport of the conditions in the participating compartments may not always be so easy as expressing the driving force as a concentration difference. Other driving forces and how to formulate them must be considered.
2. The factors that determine the value of the permeability must be investigated. Permeabilities vary according to the molecule they represent, the physical structure of the surface through which the permeant is passing, the nature of the driving force, and the fact that many biological membranes are "smart": they have permeabilities that vary according to system requirements. How to determine permeabilities, either from theories or experimental measurements, must also be considered.
When no net transport of a substance occurs between two compartments, the driving force for transport must be zero. Numerous circumstances are known in which concentration differences are finite but no net transport occurs. Several specific examples will be considered here.
Solubility effects: In a given medium, say blood plasma, with no external force applied, transport equilibrium is represented by a uniform concentration. A concentration gradient, locally, or a concentration difference between two points separated by a finite distance, is used as the driving force for transport. Each of the several, equivalent forms of Fick's law involves a flux, the diffusivity or diffusion coefficient, and a concentration gradient. You have already seen integrations of these forms that lead to the kind of permeability expression given above, involving a concentration difference between two points separated by a finite distance. However concentration is not an adequate measure of chemical potential when more than one "medium" is involved. An aqueous solution, a lipid layer serving as a membrane, a gas phase, an oil phase all are separate media and a solute distributed between two media will exhibit different concentrations in each medium -- even at equilibrium. One example: At equilibrium the concentration of oxygen in water is only 1/23,600 that of the concentration of oxygen in an ideal gas in contact with the water. (The Henry's law constant, defined as gas partial pressure in atmospheres divided by the mole fraction in the liquid of the gas at equilibrium, is 5.15 104 atm for oxygen in water at 37 C. cliquid/cgas can be shown to be 55.5/(.082 x 310 x 5.15 104).) If the solubility of oxygen in a polymer membrane placed between the aqueous phase and the gas were 10 times that of oxygen in water, one would have (on a log scale to accommodate the great differences in solubility among the media) an equilibrium distribution of oxygen like that shown in the red line, in Figure 2.
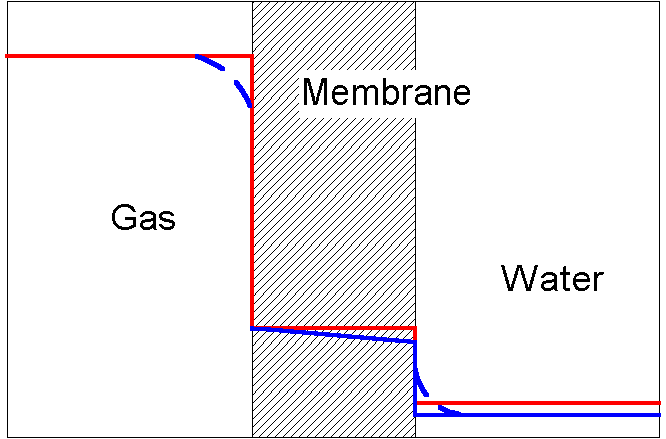
At equilibrium, there are no gradients in any phase, but there is a concentration "jump" at each interface. The concentration drops by a factor of 2360 between the gas and the membrane and by a factor of 10 between the membrane and the water phase. If there were a "sink" for oxygen in the aqueous phase, the system would not be at equilibrium and the distribution would be like that shown in the blue line, below, in which we assume that the gas and liquid media or phases are compartments, without gradients, and a linear gradient exists in the membrane. Note the discontinuity at the gas-membrane interface is not changed, because it has been assumed that the gas compartment is truly uniform and can furnish oxygen to the water phase without loss of oxygen concentration in the gas, i.e. the gas is a source. Because there is a flux in the membrane, there is a linear decrease in oxygen concentration from the gas side to the water side. Then there is a discontinuity down to a lower oxygen concentration in the water, because the sink is consuming oxygen. This lower concentration is determined by the interaction of the sink's rate of oxygen consumption and the oxygen concentration gradient that the membrane requires to deliver this much oxygen. Experiments show that equilibration at the interface occurs even when the rest of the system is not at equilibrium and finite transport is taking place.
The ratio of equilibrium concentration in one phase to that in another, reference phase is called the "partition coefficient". This number tends to be constant over a wide range of solute concentrations so long as these concentrations are such that the solute molecules interact only with membrane molecules, not with each other. Under these conditions, the solute is said to be dilute. The partition coefficient of the membrane relative to water is stated here to be 10 for this membrane with respect to oxygen.
Another, non-linear partitioning occurs often with small-molecule solutes of biological significance. One example is the distribution of oxygen between the physically dissolved component, which is the only solution component so far considered, and hemoglobin, which is present in blood. Normal blood has the capacity to bind about 20 STP ml of oxygen per 100 ml of blood when all the hemoglobin is fully saturated with oxygen. The fractional saturation, often designated as Y is related to the partial pressure of oxygen in the solution:
Y = (pO2)n / ((pO2)n + (P50)n)
where P50 is the partial pressure of oxygen that corresponds to 50% saturation. In human arterial blood, P50 is about 21 torr and n is about 2.8. The significant form of oxygen for diffusive transport is dissolved oxygen. Oxyhemoglobin diffuses slowly and does not cross biological membranes. The significant form of oxygen for convective transport is hemoglobin-bound oxygen because convection depends on the product of flow times the total content of the solute, and the hemoglobin-bound accounts for the vast majority of the releasable oxygen contained in blood. Many other substances are stored in multiple forms in blood. Metabolizable lipids are carried in the circulation by albumin, each molecule of which has multiple lipid-binding sites.
Real compartments are never uniform. They are most likely to have varying concentrations at boundaries. Near fluid boundaries, particularly solid boundaries, there is a layer not much disturbed by convection or stirring. This layer is called the "hydrodynamic boundary layer". Within (sometimes beyond) this layer is a layer of fluid in which exist concentration gradients of molecules being transported. This layer is called the "concentration boundary layer". In small systems, such as cells, these layers are negligible. In larger systems such as arteries and veins, and in artificial organs, these layers are larger and may add further resistance to mass transfer. The dashed blue lines in Figure 2 show the effect of a concentration boundary layer on the concentration profile during transport.
The effect of fixed charge (the Donnan effect): In many biological compartments one has ionized solutes, only some of which can diffuse across the compartmental boundary. Consider a dissolved and impermeable protein anion with a charge of Z negative charges per molecule. The only accompanying ions are Na+ and Cl-, both permeable. Equilibrium and thus no net transport is achieved when the free energy of all species to which the membrane is permeable is the same on each side of the membrane. If a membrane separates two compartments, 1 and 2, the equivalent equilibrium condition in dilute solution is that [Na+] [Cl-]|1 = [Na+] [Cl-]|2. This condition must be met without violating the requirement of charge equality. If x1 moles of Na Z P are added to compartment 1 and x2 moles of NaCl are added to compartment 2, y moles of Na+ and Cl- can be presumed to move from compartment 2 to compartment 1. The equilibrium condition is
(Zx1 + y)(y) = (x2 - y) (x2 - y), or y = x22/(Zx1 + 2x2)
from which one can calculate all the equilibrium conditions, given an initial distribution of ions. Blood plasma might (as a gross simplification) be presumed to be 6 10-4 M in albumin and 0.15 M in NaCl. Albumin molecules might have a Z of about 8 at the body pH of 7.4. Then the ratio of chloride ion between a solution of 0.15 M NaCl and a solution of Na8 albumin would be about 0.97.
Both of the foregoing examples show that when external forces are not present, driving forces should be measured as departures from an equilibrium condition that is not always expressible as equality of concentration between two compartments.
Transport in an electrical potential gradient: When an external force is applied to a system the equilibrium condition is changed. While charged molecules (ions) are the elements of a solution that respond directly to a potential difference between two compartments, their movement affects many other cellular processes. The interaction between the electric potential gradient and charged species is formulated as a flux that acts in tandem with the Fickian diffusion flux. For the i th charged species:
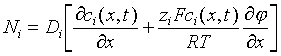
and, to obtain the current density:
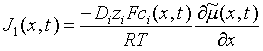
potential related to the movement of species i is defined as
![]()
![]()
This quantity, when differentiated, and substituted into the preceding equation produces an expression for Ji equal to the right-hand side of the expression for Ni, multiplied by ziF, as expected. The expression for Ji is one form of the Nernst-Planck equation. The quantity m i0 is the reference value of the electrochemical potential, evaluated at unit concentration and zero potential. The respective terms that depend on solute concentration and potential are the stored chemical energy and the stored electrostatic energy per mole. (Weiss, v. 1, p. 456, 1996) Thus, the flux of ions and the current density carried by the ions are proportional to the electrochemical potential gradient. Equilibrium, construed as a zero net flux of ions, is given by the relationship
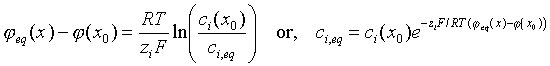
Arguments can be made that support well the general assumption that electroneutrality must always be expected: one cannot change the accumulation of charge in any finite volume over any finite time. (Weiss, section 7.2.3.) The mathematically equivalent statement is that the divergence of the sum of all ion current densities is identically zero.
The Nernst-Planck equation can be integrated to give an equation for steady-state transport of ions through a homogeneous membrane of thickness t .
Because Ji is invariant through the membrane, the equation can be integrated thus:
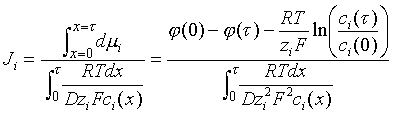
The ci in this equation are intramembrane concentrations. If however they are proportional to concentrations ci and co in the adjacent liquids with the same partition coefficient, the ratio in the logarithm can be replaced by the corresponding ratio of liquid concentrations. One can then write:
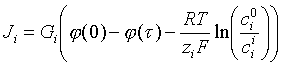
the logarithmic term representing the Nernst equilibrium potential difference across the membrane. When the measured potential difference equals the opposite of the Nernst equilibrium potential difference, which is caused by a concentration ratio, as shown, there is no transport. The coefficient Gi which functions like a permeability is called the membrane conductance. The same form of equation can be applied to a heterogeneous membrane (in which diffusion occurs through pores or channels, frequently "gated" by a biological signal molecule). In these cases, the only difference is in the form of the expression for the conductance.
Since the Nernst equilibrium potential is measurable and important in determining the flow of ions, its magnitude is of interest. A calculation taught to every medical student, and worth memorizing, is that a 10-fold change in the concentration of a monovalent metabolite across a membrane gives rise to a transmembrane voltage of 59 millivolts. Since only one potential difference can exist across a membrane, once the value of this potential difference is fixed the equilibrium ratio of concentrations for all ions across that membrane is also fixed.
The movement of water across cell membranes is, at root, no different from the passive movement of other small molecules. However, rather than reckon a water concentration, one calculates something close to the molar concentration of all the impermeant molecules in the solution and then multiplies this summed concentration by RT to obtain the osmotic pressure (the vantHoff relation, valid in dilute solution, c = P/RT). Since the osmotic pressure is formed from a complement of the water concentration, it is not surprising that water moves in the direction of increasing osmotic pressure. What makes water transport calculations different is the high permeability of water and the very high concentration (55 moles/L) normally associated with it. Osmotic pressure can be offset by hydrostatic pressure acting in the same direction. Osmotic equilibrium is achieved when the hydrostatic and osmotic pressures are equal and the driving force for water transport in systems capable of maintaining a pressure gradient must be formed as a deviation from this equilibrium: NH2O = LP (D P - D P ). It is important to remind oneself that water movement is not caused by pressure, but rather by pressure difference. Thus pressuring a system will not generally cause water movement unless some stress-bearing membranes support a pressure difference between different parts of the system. It is also important to appreciate the difference between the osmotic pressure and the oncotic component of this pressure. Osmotic pressure can be caused by diffusible molecules to the extent that a given membrane can distinguish them from water. For diffusible molecules the osmotic pressure contribution is written as s iciRT, where s i is the "reflection coefficient," equal to zero for a molecule indistinguishable by the membrane from water and equal to unity for a non-permeant molecule. The value of s i will vary between zero and unity for diffusible molecules and the concentration of these molecules will fall over time as they move across the membrane, decreasing the net concentration difference. Non-diffusible molecules, such as albumin in blood plasma, generate an oncotic pressure that is maintained indefinitely. Note that the value of s i depends on both the solute molecule and the membrane. A proper calculation of the osmotic driving force in a multi-solute system is RTS is ici.
Active and facilitated transport: This discussion has related the rate of movement of molecules across a compartment boundary or membrane to a permeability multiplied by a driving force, where the driving force is a departure of a concentration or potential from its equilibrium value. As Weiss notes, the natural dissipation of energy associated with transport of electrons and molecules, the practical inability of all species to be in Nernstian equilibrium across a membrane that can support only one potential difference, and the question of how concentration gradients could have been established in the first place -- all suggest that the story of transport across membranes involve more than the flow of molecules down a concentration gradient. The additional factor is the actual pumping of molecules and ions against (or "up") an electrochemical potential gradient, called active transport.
Active transport may be either electrogenic, each cycle of the pump transfers charge, or nonelectrogenic: either a neutral molecule or counterbalancing charged molecules are transferred. Examples of the latter include the transfer of a sodium ion in one direction concomitant with the transfer of a potassium ion in the opposite direction, or the transfer of an anion and cation, or a neutral molecule in one direction. Active transport requires the provision of free energy because the movement of a molecule up an electrochemical concentration gradient, or from a compartment at one level of electrochemical potential to another at a higher electrochemical potential, causes it to gain free energy and this free energy must be gotten from another molecule, usually one of the high-energy molecules used by the cell, e.g. ATP. The best studied of the ion pumps is that responsible for the exchange of sodium and potassium. This antiport pump is essential to the maintenance of the internal environment of mammalian cells. Paraphrasing Weiss who quotes Lew and Bookchin (1986), the pump rate can be represented as:
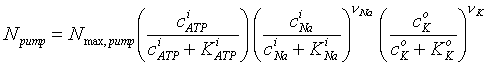
One sees that the pump depends in a saturable manner on all three reactants: intracellular ATP and sodium, and extracellular potassium. In this model, the pump behaves as an irreversible enzyme reaction involving three substituents. In fact, there is some effect of the product concentrations, extracellular sodium and intracellular potassium. The pump has been studied carefully and the ratio of sodium efflux to potassium influx has been shown to be about 3:2, with 1 molecule of ATP hydrolyzed for each 3 moles of sodium pumped. Thus, in the formula above, n Na is to be taken as 3 and n K as 2. There are between 500 and 5000 of these pumps/m m2 in typical nerve, muscle, kidney and heart cells.
Cells have a complex internal structure including organelles many of which have their own membranes and many questions that relate to transport within cells are still being actively addressed. Here is considered only transport across the plasma membrane. Much of the material is taken from the first edition of Molecular Cell Biology, Darnell, J., Lodisch, H., and Baltimore, D., (1988). Comparable texts, even those published much more recently, have very similar treatments.
The common structural element in the membranes of all cells is a phospholipid bilayer, whose thickness is about 6 nm. This layer can be seen as a thin fluid layer crowded with floating, moving protein molecules. Most of these proteins pass clear through the membrane and thus have cytoplasmic, extracelluar, and transmembrane regions, with the carboxy terminus usually in the cytoplasm and the amino terminus in the extracellular fluid. Protein molecules are linear and many of them make multiple passes through the membrane, almost as does a thread when used to sew a button to a piece of clothing. Some cells, particularly those, which adhere to each other to form a macroscopic barrier, have a different membrane composition on opposite sides of the cell. Examples include the endothelial lining of blood vessels and the intestinal epithelium. Transport of molecules through the "gap" junctions that exist between adjacent cells in the barrier layer is important.
A pure, artificial phospholipid bilayer is permeable to water, to small hydrophobic molecules (including the dissolved gasses carbon dioxide, nitrogen, oxygen, and halogen anesthetics) and to small uncharged polar molecules such as urea and ethanol. It is impermeable to large, uncharged polar molecules such as glucose, to ions, and to charged polar molecules such as amino acids, ATP, and glucose 6-phosphate. Permeability to small molecules depends on the oil-water partition coefficient. Glycerol has a permeability (cm/h) of 0.008 and a partition coefficient of about 0.00008. Trimethycitrate has a permeability of 0.8 and a partition coefficient of about 0.08.
Many molecules are transported by permeases. This kind of transport is saturable: as with enzymes catalyzing chemical reactions, the rate of transport depends on permeant concentration on the "source" side of the membrane but the dependence decreases with increasing concentration. A maximum rate of transport is approached asymptotically, as determined by a Michaelis-Menten expression and the value of a KM (with units of concentration) in the denominator. Permeases may effect either facilitated transport, in which transport occurs "down" a concentration gradient, or active transport, in which transport may occur "up" or against a concentration gradient. With facilitated transport, the free energy change associated with the transport is generally small, perhaps zero, and it is important to use enzyme kinetics, which allow for transport in both directions, i.e. reversibility. Sometimes facilitated transport of ions is coordinated. Band three protein causes the one-for-one, nonelectrogenic exchange of chloride and bicarbonate. In systemic capillaries carbon dioxide generated in the tissues by oxidation of glucose permeates the lipid bilayer and enters red blood cells as they pass through the capillaries. Carbonic anhydrase, which is concentrated in the red blood cells, catalyzes the hydrolysis of CO2 to carbonic acid, forming hydrogen ions, which stay in the cell, buffered by hemoglobin, and HCO3- ions which are exchanged for Cl- from the plasma. In the lungs the process is reversed, leading to the formation of CO2 which permeates the lipid bilayer and is blown off with expired air.
With active transport the overall free energy change associated with the transport is generally large and the reaction may often be assumed irreversible. (See the discussion of, and model for, the Na-K pump, above.) In this case, as with enzyme reactions that use high-energy molecules, the positive free energy change of the desired reaction is offset by free energy taken from the high-energy molecule. The only substances known to be actively transported by ATP-driven pumps are Na+, K+, H+, and Ca++. You will learn about the importance of these pumps at both the cellular and organismic level in your biology course. However, it is crucial to comprehend how pumps that move only these four cations can cause many other substances against electrochemical gradients. All additional transport of vital molecules is caused by cotransport: the processes of symport (same direction) and antiport (opposite directions). The process of uncoupled transport using permeases is referred to as uniport.
Permeases are translational catalysts in the sense that they cause the movement (translation) rather than the chemical transformation of their substrates. As with catalysts, reactions involving permeases are reversible. The mitochondrion is the organelle in which is formed most of the ATP generated in a cell capable of oxidative metabolism. The flow of hydrogen ion down a concentration gradient provides the energy for an ATP-ADP translocase that antiports an ADP molecule into, and concurrently an ATP molecule out of, the mitochondrion, assuring the high concentration of ADP and the low concentration of ATP within the mitochondrion necessary to drive the reaction toward the formation of ATP.
A specific glucose-Na+ symport protein located in the microvillar membranes accomplishes the movement of glucose from the intestinal lumen across the apical surface of the brush border cells of the intestinal wall. The transport of one glucose molecule is obligatorily coupled to that of one Na+ ion. The process proceeds because the concentration of Na+ is much higher in the lumen than in the cell. It should be obvious, however, that the process can only be maintained if the symported Na+ is somehow pumped out. The Na-K-ATPase pump accomplishes the pumping. Cardiac muscle cells use an antiport protein to move calcium out of the cell in exchange for a sodium ion moved into the cell.
The movement of very large molecules and micro and nanoparticles to and from cells will not be discussed here in any detail. The process of endocytosis involves the adsorption or binding of the entity to be transported to the cell surface and the stepwise enclosure of the adsorbed entity by cell membrane. Eventually a droplet of cell membrane enclosing the entity appears in the cytoplasm and is then processed. The membrane material, and possibly the ingested entity, is, after reprocessing, recycled to the surface. The reverse process of exocytosis results in the secretion of macromolecules from within the cell.
Return to course main page.